

cNap1 bridges centriole contact sites to maintain centrosome cohesion
- PMID: 36282799
- PMCID: PMC9595518
- DOI: 10.1371/journal.pbio.3001854
cNap1 bridges centriole contact sites to maintain centrosome cohesion
Abstract
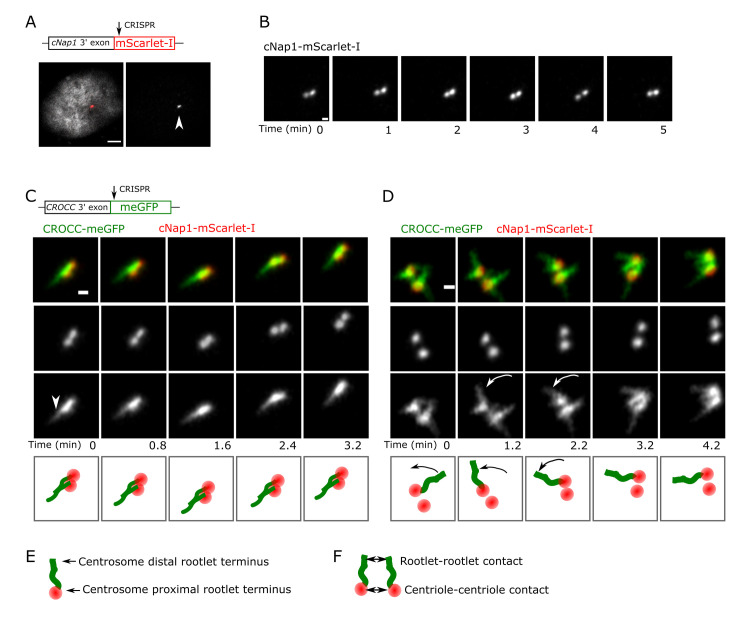
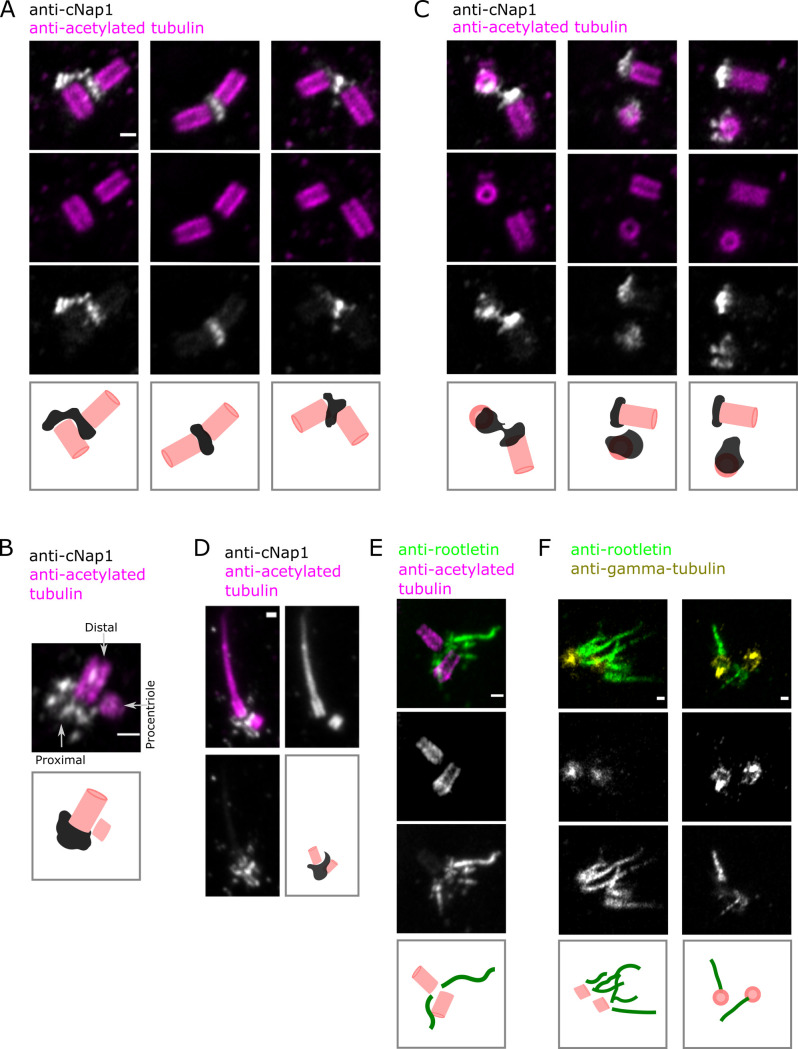
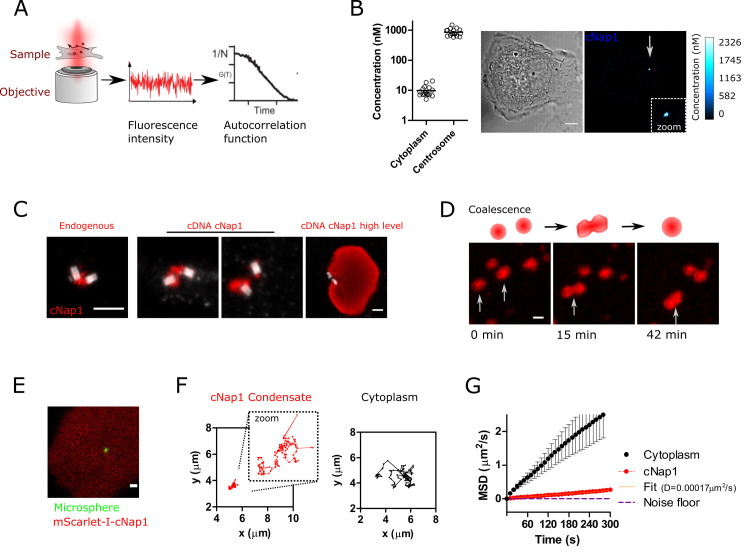
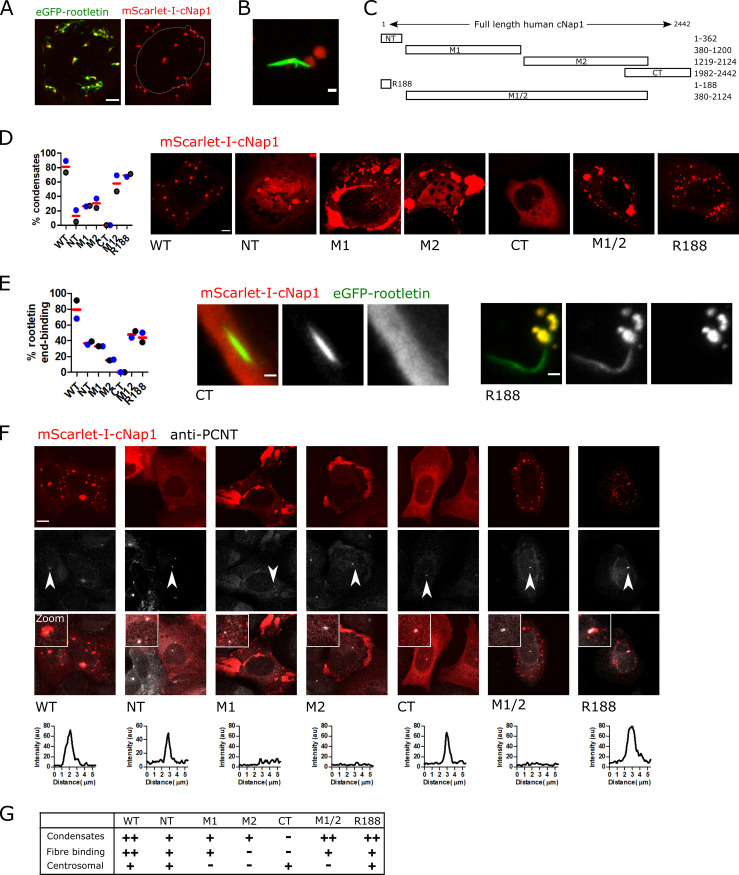
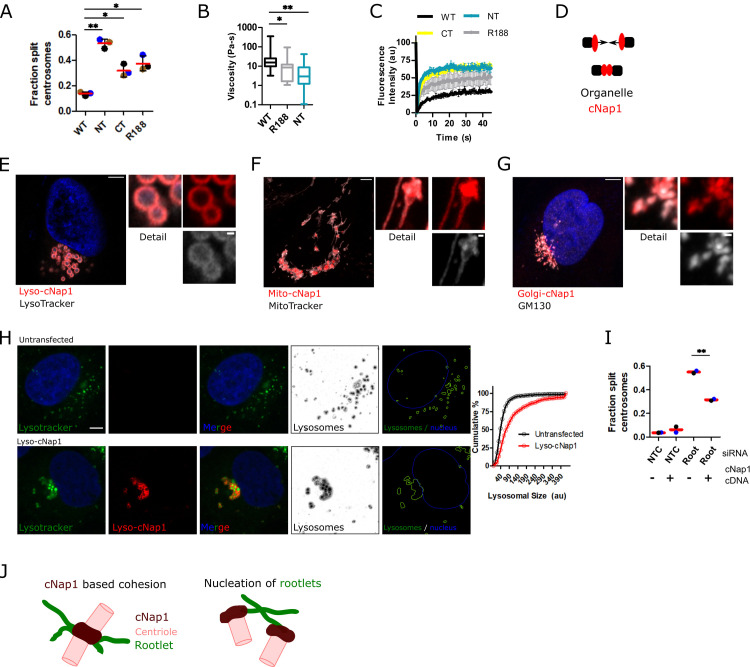
Centrioles are non-membrane-bound organelles that participate in fundamental cellular processes through their ability to form physical contacts with other structures. During interphase, two mature centrioles can associate to form a single centrosome-a phenomenon known as centrosome cohesion. Centrosome cohesion is important for processes such as cell migration, and yet how it is maintained is unclear. Current models indicate that pericentriolar fibres termed rootlets, also known as the centrosome linker, entangle to maintain centriole proximity. Here, I uncover a centriole-centriole contact site and mechanism of centrosome cohesion based on coalescence of the proximal centriole component cNap1. Using live-cell imaging of endogenously tagged cNap1, I show that proximal centrioles form dynamic contacts in response to physical force from the cytoskeleton. Expansion microscopy reveals that cNap1 bridges between these contact sites, physically linking proximal centrioles on the nanoscale. Fluorescence correlation spectroscopy (FCS)-calibrated imaging shows that cNap1 accumulates at nearly micromolar concentrations on proximal centrioles, corresponding to a few hundred protein copy numbers. When ectopically tethered to organelles such as lysosomes, cNap1 forms viscous and cohesive assemblies that promote organelle spatial proximity. These results suggest a mechanism of centrosome cohesion by cNap1 at the proximal centriole and illustrate how a non-membrane-bound organelle forms organelle contact sites.
Conflict of interest statement
The author has declared that no competing interests exist.
Figures
Update of
-
Stable centrosomal roots disentangle to allow interphase centriole independence.PLoS Biol. 2018 Apr 12;16(4):e2003998. doi: 10.1371/journal.pbio.2003998. eCollection 2018 Apr. PLoS Biol. 2018. Update in: PLoS Biol. 2022 Oct 25;20(10):e3001854. doi: 10.1371/journal.pbio.3001854. PMID: 29649211 Free PMC article. Updated.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
