

Phosphorylation of RXRα mediates the effect of JNK to suppress hepatic FGF21 expression and promote metabolic syndrome
- PMID: 36282921
- PMCID: PMC9636906
- DOI: 10.1073/pnas.2210434119
Phosphorylation of RXRα mediates the effect of JNK to suppress hepatic FGF21 expression and promote metabolic syndrome
Abstract
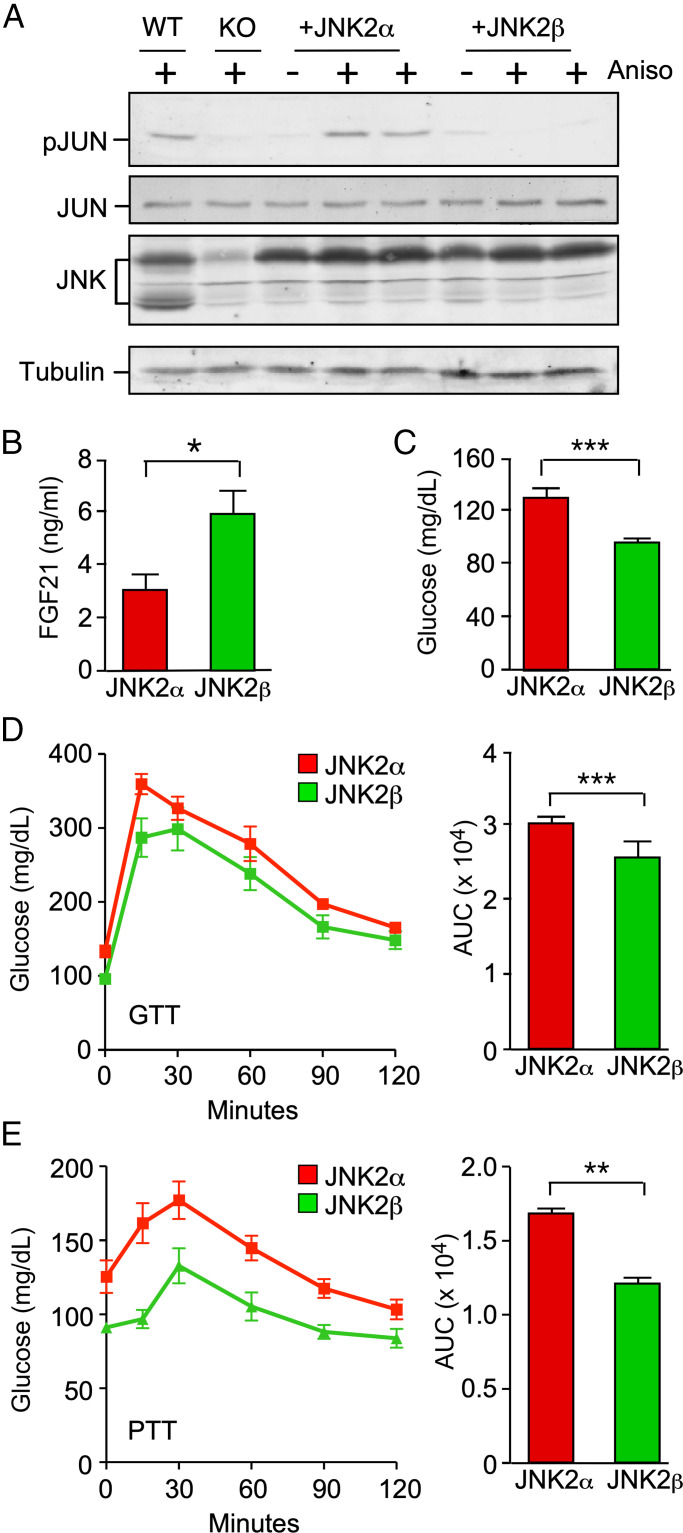
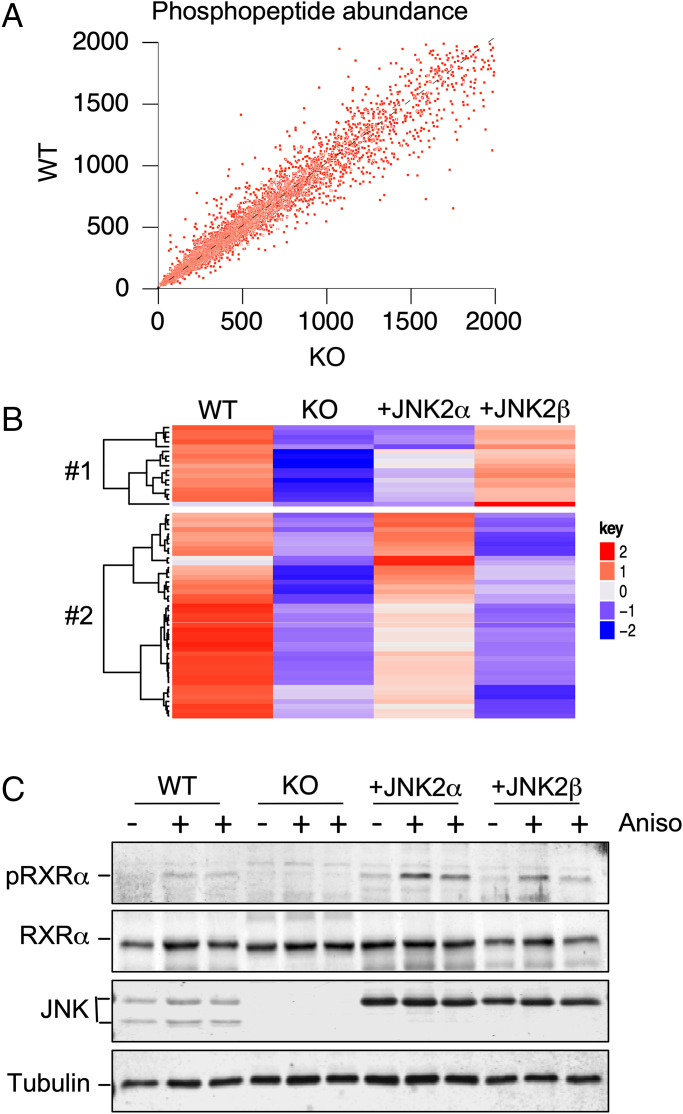
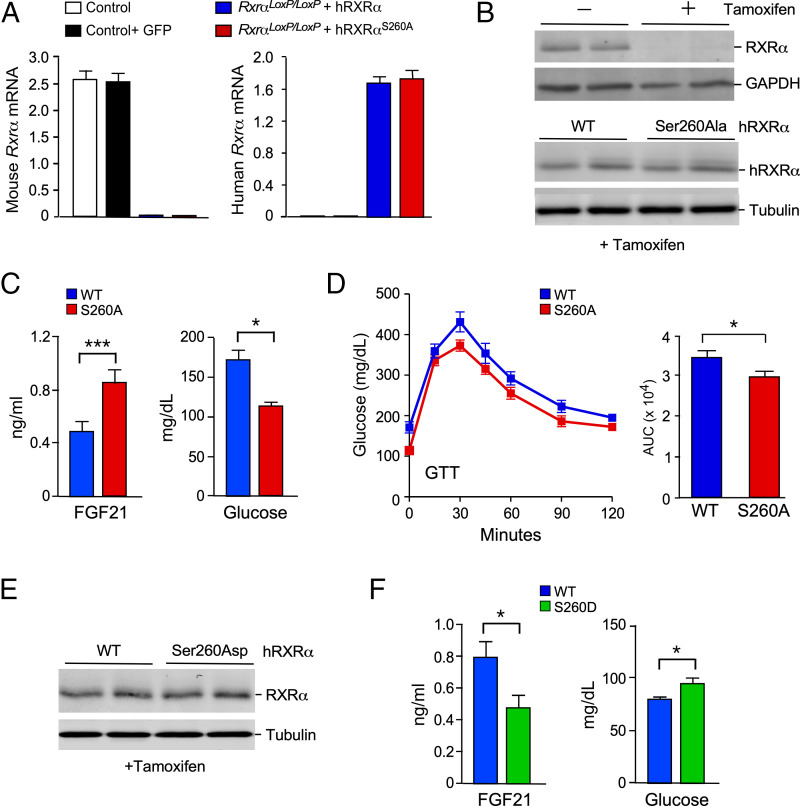
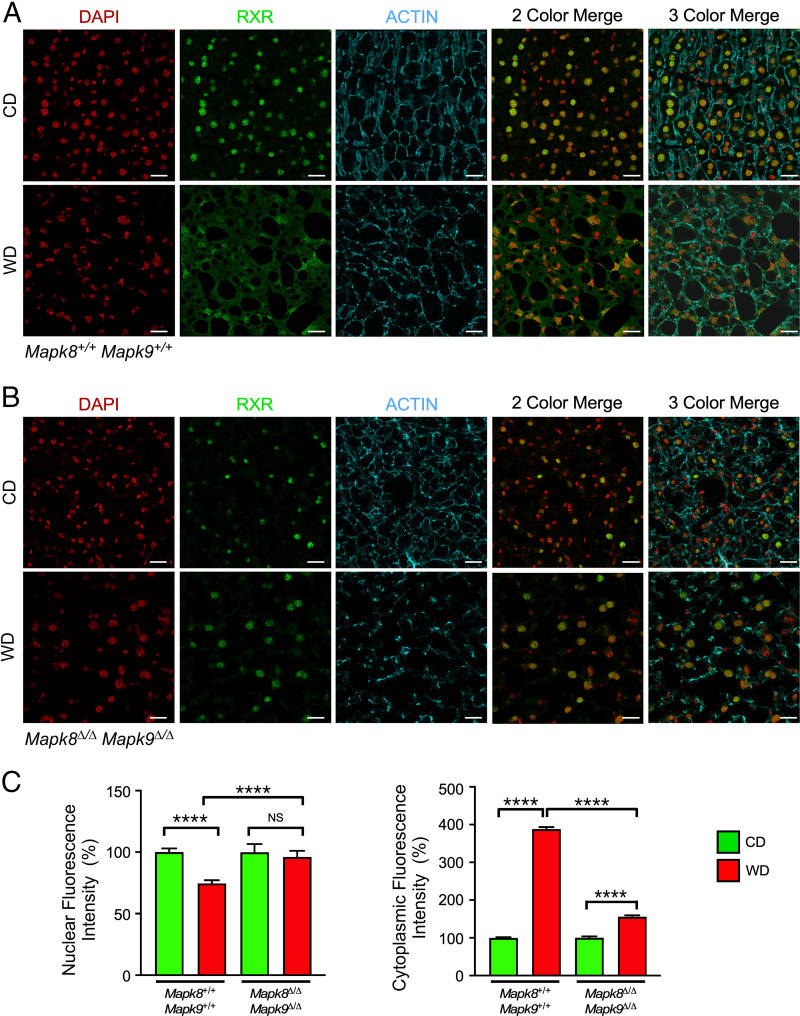
The cJun NH2-terminal kinase (JNK) signaling pathway in the liver promotes systemic changes in metabolism by regulating peroxisome proliferator-activated receptor α (PPARα)-dependent expression of the hepatokine fibroblast growth factor 21 (FGF21). Hepatocyte-specific gene ablation studies demonstrated that the Mapk9 gene (encoding JNK2) plays a key mechanistic role. Mutually exclusive inclusion of exons 7a and 7b yields expression of the isoforms JNK2α and JNK2β. Here we demonstrate that Fgf21 gene expression and metabolic regulation are primarily regulated by the JNK2α isoform. To identify relevant substrates of JNK2α, we performed a quantitative phosphoproteomic study of livers isolated from control mice, mice with JNK deficiency in hepatocytes, and mice that express only JNK2α or JNK2β in hepatocytes. We identified the JNK substrate retinoid X receptor α (RXRα) as a protein that exhibited JNK2α-promoted phosphorylation in vivo. RXRα functions as a heterodimeric partner of PPARα and may therefore mediate the effects of JNK2α signaling on Fgf21 expression. To test this hypothesis, we established mice with hepatocyte-specific expression of wild-type or mutated RXRα proteins. We found that the RXRα phosphorylation site Ser260 was required for suppression of Fgf21 gene expression. Collectively, these data establish a JNK-mediated signaling pathway that regulates hepatic Fgf21 expression.
Keywords: FGF21; JNK; RXRα; high-fat diet; insulin resistance.
Conflict of interest statement
The authors declare no competing interest.
Figures
References
-
- Badman M. K., et al. , Hepatic fibroblast growth factor 21 is regulated by PPARalpha and is a key mediator of hepatic lipid metabolism in ketotic states. Cell Metab. 5, 426–437 (2007). - PubMed
-
- Inagaki T., et al. , Endocrine regulation of the fasting response by PPARalpha-mediated induction of fibroblast growth factor 21. Cell Metab. 5, 415–425 (2007). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
