

Inhibition of the expression of rgs-3 alleviates propofol-induced decline in learning and memory in Caenorhabditis elegans
- PMID: 36284438
- PMCID: PMC9804065
- DOI: 10.1111/cns.14004
Inhibition of the expression of rgs-3 alleviates propofol-induced decline in learning and memory in Caenorhabditis elegans
Abstract
Background: Exposure to anesthesia leads to extensive neurodegeneration and long-term cognitive deficits in the developing brain. Caenorhabditis elegans also shows persistent behavioral changes during development after exposure to anesthetics. Clinical and rodent studies have confirmed that altered expression of the regulators of G protein signaling (RGS) in the nervous system is a factor contributing to neurodegenerative and psychological diseases. Evidence from preclinical studies has suggested that RGS controls drug-induced plasticity, including morphine tolerance and addiction. This study aimed to observe the effect of propofol exposure in the neurodevelopmental stage on learning and memory in the L4 stage and to study whether this effect is related to changes in rgs-3 expression.
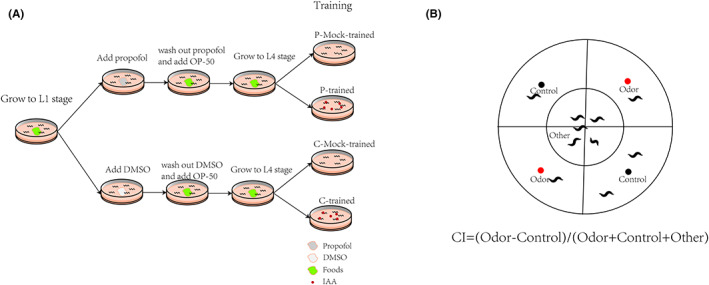
Methods: Caenorhabditis elegans were exposed to propofol at the L1 stage, and learning and memory abilities were observed at the L4 stage. The expression of rgs-3 and the nuclear distribution of EGL-4 were determined to study the relevant mechanisms. Finally, RNA interference was performed on rgs-3-expressing cells after propofol exposure. Then, we observed their learning and memory abilities.
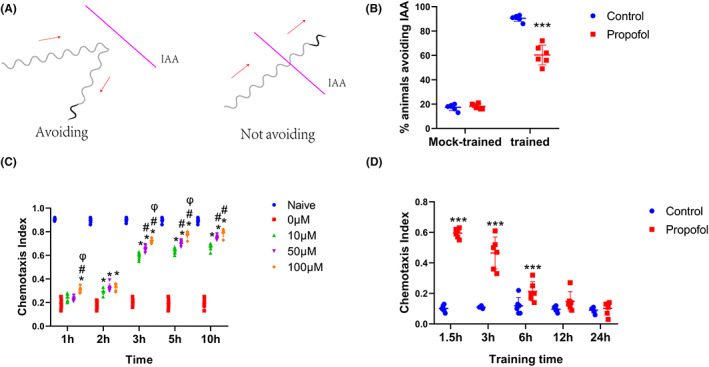
Results: Propofol time- and dose-dependently impaired the learning capacity. Propofol induced a decline in non-associative and associative long-term memory, rgs-3 upregulation, and a failure of nuclear accumulation of EGL-4/PKG in AWC neurons. Inhibition of rgs-3 could alleviate the propofol-induced changes.
Conclusion: Inhibition of the expression of rgs-3 alleviated propofol-induced learning and memory deficits in Caenorhabditis elegans.
Keywords: learning; memory; neurodevelopment; propofol; regulators of G protein signaling.
© 2022 The Authors. CNS Neuroscience & Therapeutics published by John Wiley & Sons Ltd.
Conflict of interest statement
The authors declare no conflicting financial interest.
Figures




References
-
- Gundacker C, Forsthuber M, Szigeti T, et al. Lead (Pb) and neurodevelopment: A review on exposure and biomarkers of effect (BDNF, HDL) and susceptibility. Int J Hyg Environ Health. 2021;238:113855. - PubMed
-
- Nakaoka H, Hisada A, Matsuzawa D, et al. Associations between prenatal exposure to volatile organic compounds and neurodevelopment in 12‐month‐old children: the Japan Environment and Children's Study (JECS). Sci Total Environ. 2021;794:148643. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
