

Mechanisms of Action of the Peptide Toxins Targeting Human and Rodent Acid-Sensing Ion Channels and Relevance to Their In Vivo Analgesic Effects
- PMID: 36287977
- PMCID: PMC9612379
- DOI: 10.3390/toxins14100709
Mechanisms of Action of the Peptide Toxins Targeting Human and Rodent Acid-Sensing Ion Channels and Relevance to Their In Vivo Analgesic Effects
Abstract
Acid-sensing ion channels (ASICs) are voltage-independent H+-gated cation channels largely expressed in the nervous system of rodents and humans. At least six isoforms (ASIC1a, 1b, 2a, 2b, 3 and 4) associate into homotrimers or heterotrimers to form functional channels with highly pH-dependent gating properties. This review provides an update on the pharmacological profiles of animal peptide toxins targeting ASICs, including PcTx1 from tarantula and related spider toxins, APETx2 and APETx-like peptides from sea anemone, and mambalgin from snake, as well as the dimeric protein snake toxin MitTx that have all been instrumental to understanding the structure and the pH-dependent gating of rodent and human cloned ASICs and to study the physiological and pathological roles of native ASICs in vitro and in vivo. ASICs are expressed all along the pain pathways and the pharmacological data clearly support a role for these channels in pain. ASIC-targeting peptide toxins interfere with ASIC gating by complex and pH-dependent mechanisms sometimes leading to opposite effects. However, these dual pH-dependent effects of ASIC-inhibiting toxins (PcTx1, mambalgin and APETx2) are fully compatible with, and even support, their analgesic effects in vivo, both in the central and the peripheral nervous system, as well as potential effects in humans.
Keywords: APETx2; ASIC; MitTx; PcTx1; mambalgin; nociception; pain; peptide; sodium channels; toxins.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
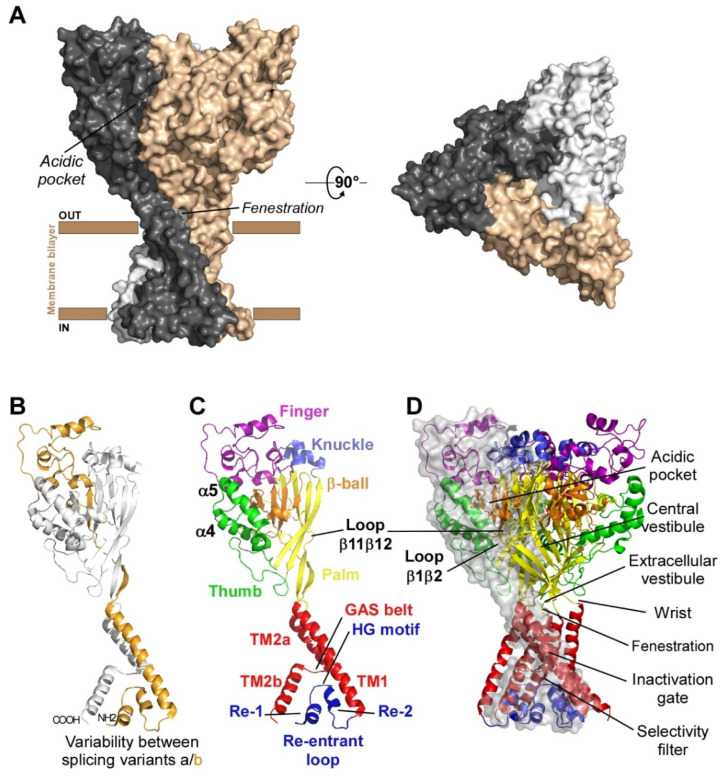
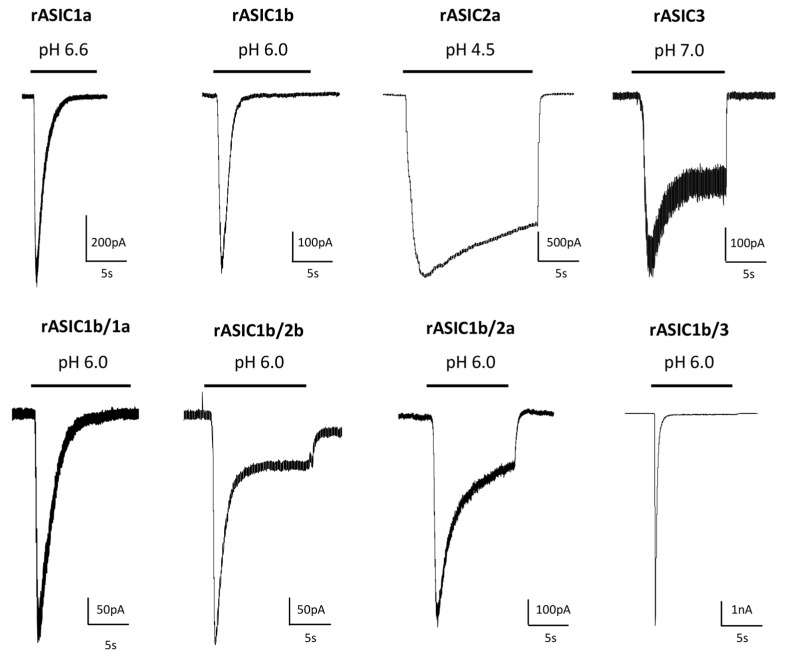
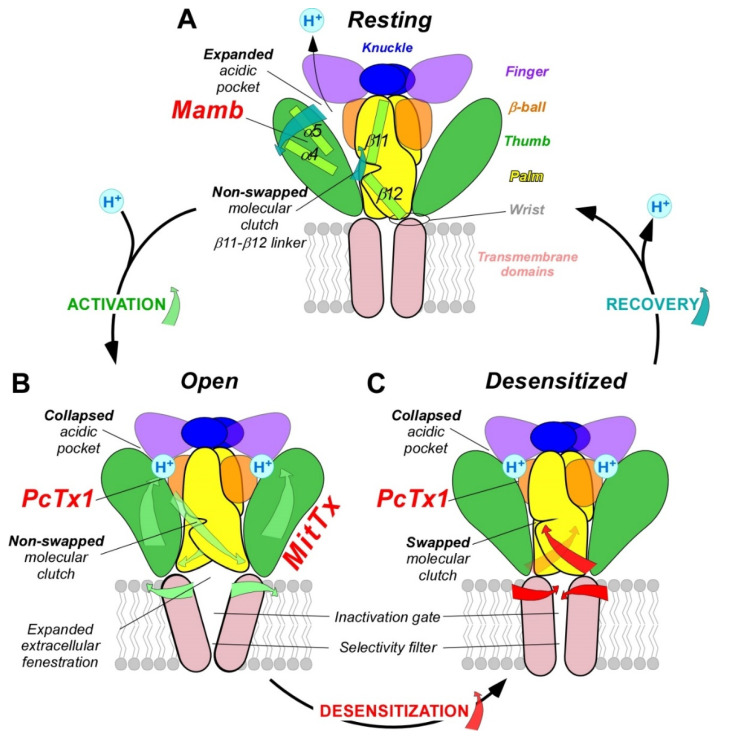
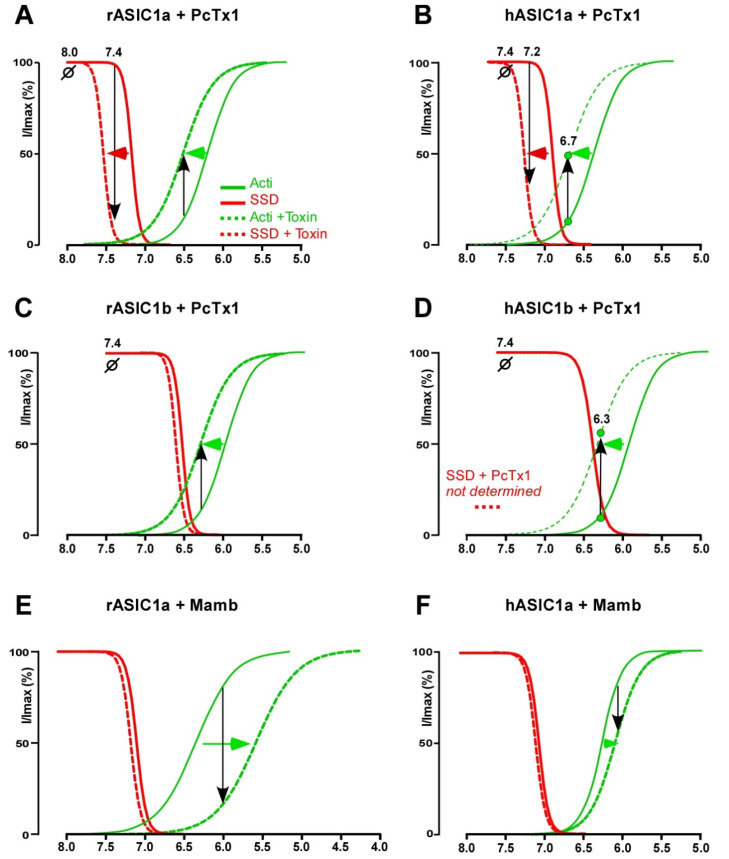
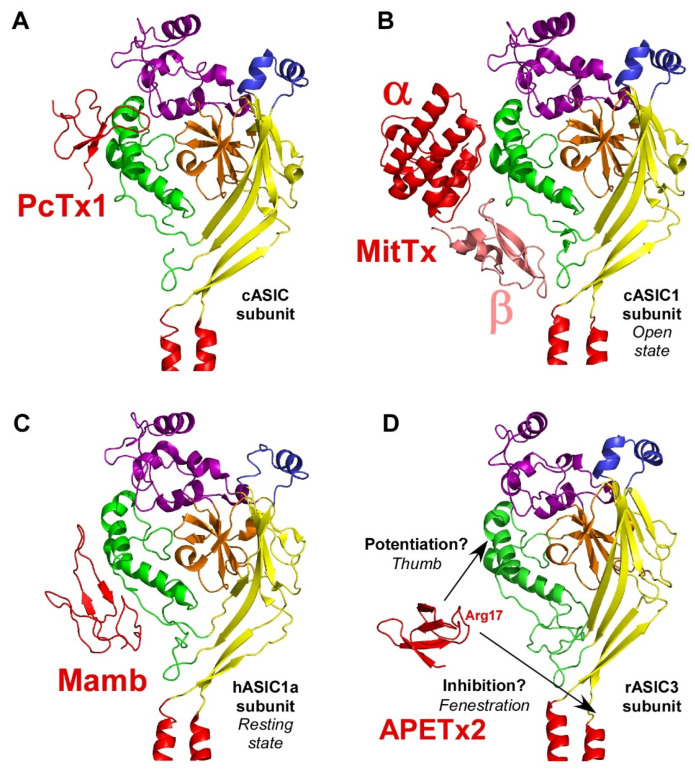
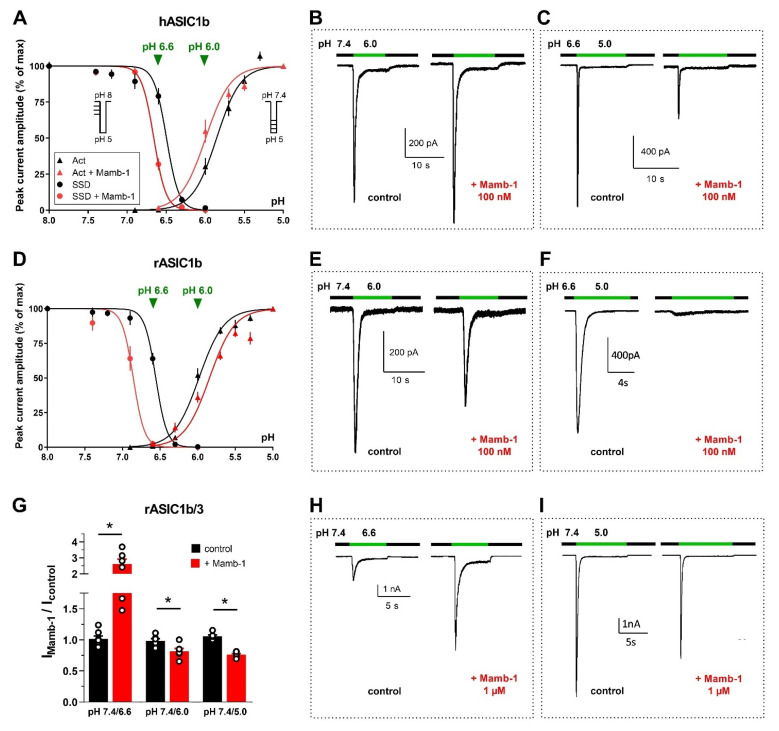
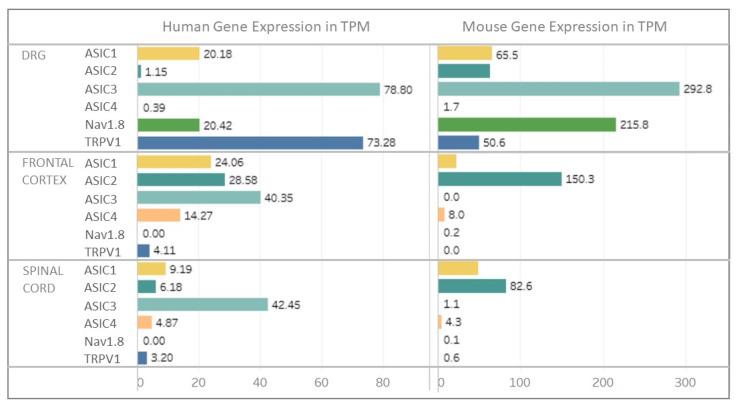
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
