

Clonal expansion and epigenetic inheritance of long-lasting NK cell memory
- PMID: 36289449
- PMCID: PMC9663309
- DOI: 10.1038/s41590-022-01327-7
Clonal expansion and epigenetic inheritance of long-lasting NK cell memory
Abstract
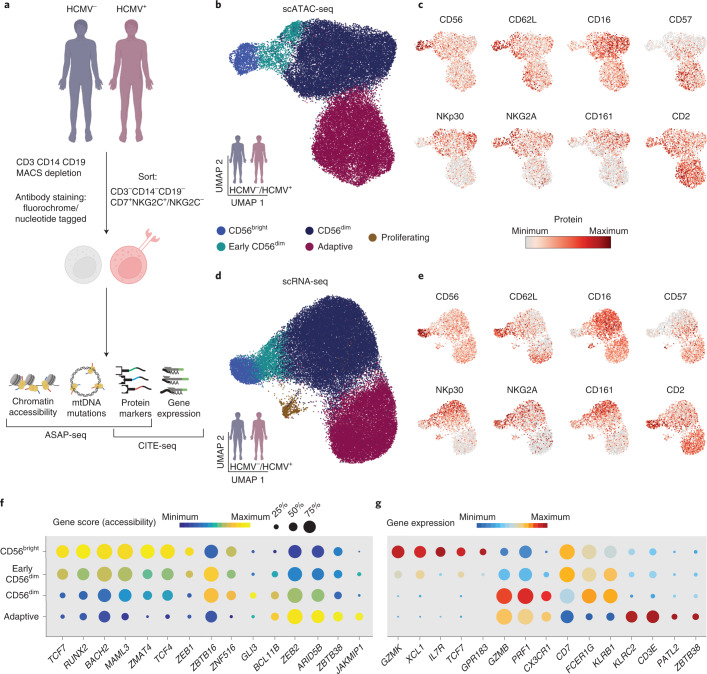
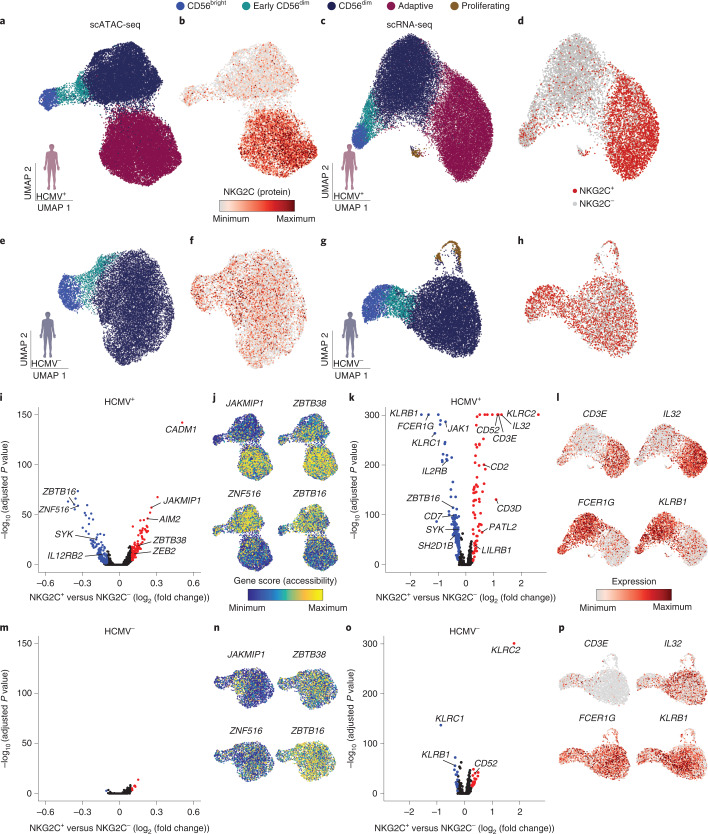
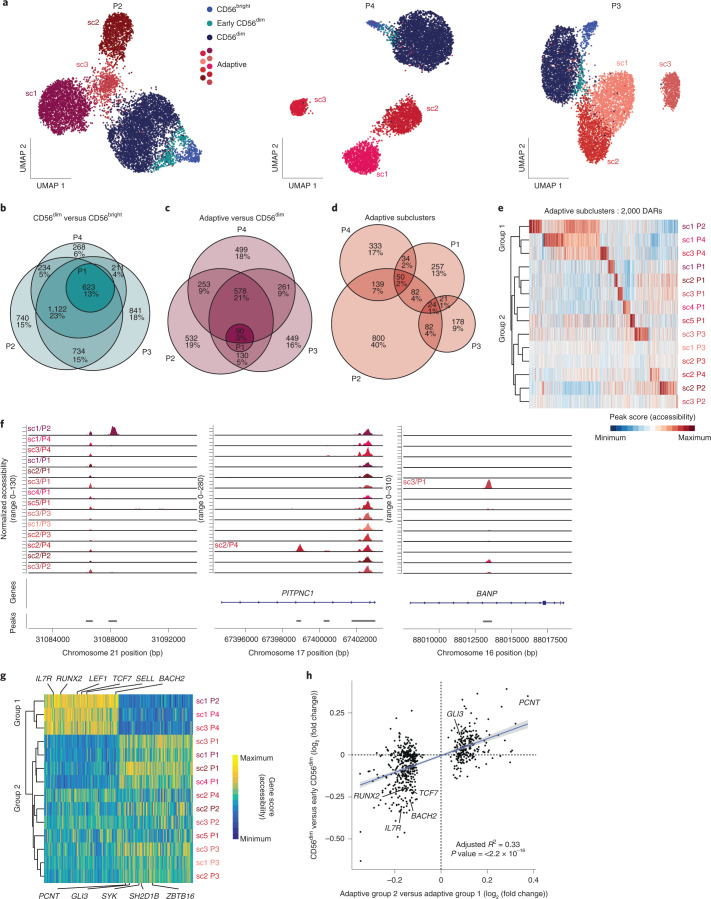
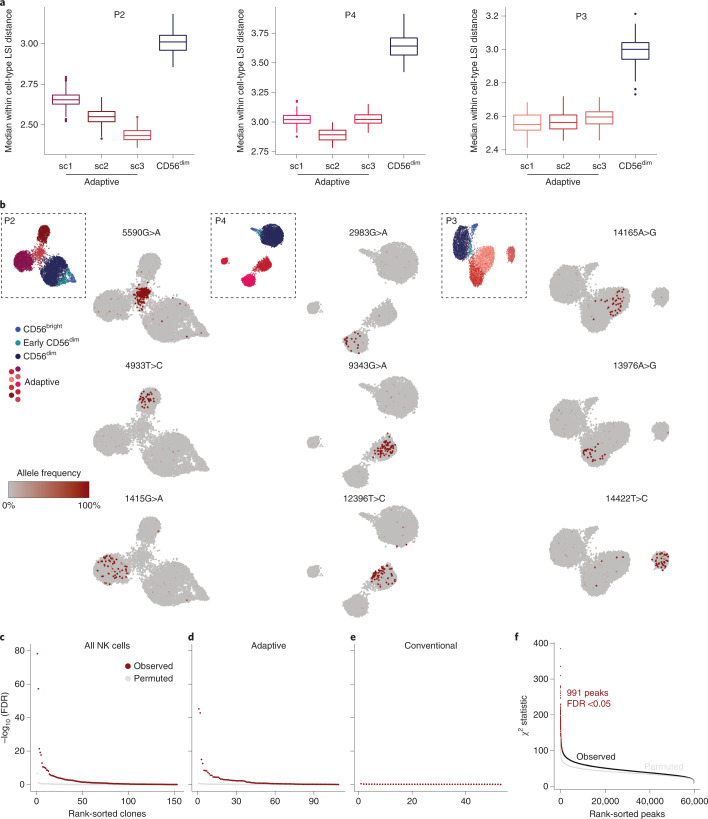
Clonal expansion of cells with somatically diversified receptors and their long-term maintenance as memory cells is a hallmark of adaptive immunity. Here, we studied pathogen-specific adaptation within the innate immune system, tracking natural killer (NK) cell memory to human cytomegalovirus (HCMV) infection. Leveraging single-cell multiomic maps of ex vivo NK cells and somatic mitochondrial DNA mutations as endogenous barcodes, we reveal substantial clonal expansion of adaptive NK cells in HCMV+ individuals. NK cell clonotypes were characterized by a convergent inflammatory memory signature enriched for AP1 motifs superimposed on a private set of clone-specific accessible chromatin regions. NK cell clones were stably maintained in specific epigenetic states over time, revealing that clonal inheritance of chromatin accessibility shapes the epigenetic memory repertoire. Together, we identify clonal expansion and persistence within the human innate immune system, suggesting that these mechanisms have evolved independent of antigen-receptor diversification.
© 2022. The Author(s).
Conflict of interest statement
T.R. and C.R. are listed as inventors on a patent series covering NKG2C-activating peptides. The other authors declare no competing interests.
Figures












Comment in
-
Thanks for the NK cell memories.Nat Immunol. 2022 Nov;23(11):1512-1514. doi: 10.1038/s41590-022-01336-6. Nat Immunol. 2022. PMID: 36289448 No abstract available.
-
Bearing a grudge-How human natural killer cells remember past insults.Sci Immunol. 2023 Jan 6;8(79):eadg4688. doi: 10.1126/sciimmunol.adg4688. Epub 2023 Jan 6. Sci Immunol. 2023. PMID: 36608111
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
