

The C-Terminus of the PSMA3 Proteasome Subunit Preferentially Traps Intrinsically Disordered Proteins for Degradation
- PMID: 36291102
- PMCID: PMC9600399
- DOI: 10.3390/cells11203231
The C-Terminus of the PSMA3 Proteasome Subunit Preferentially Traps Intrinsically Disordered Proteins for Degradation
Abstract
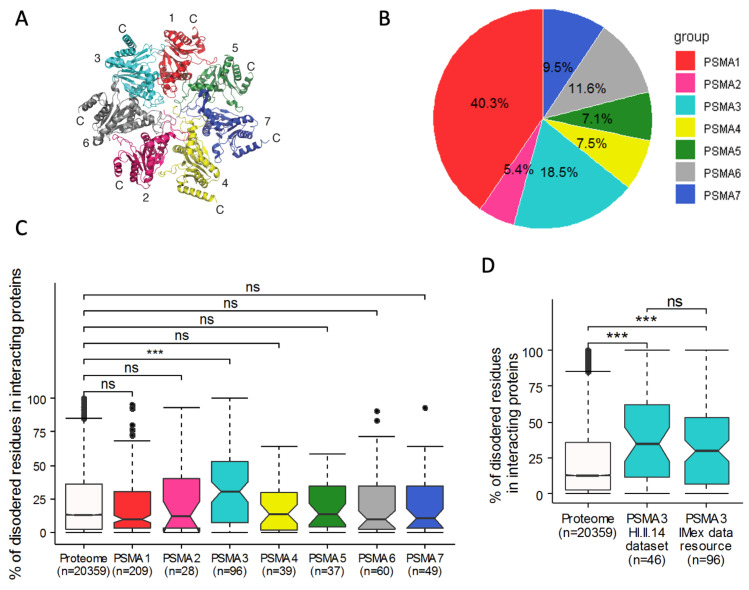
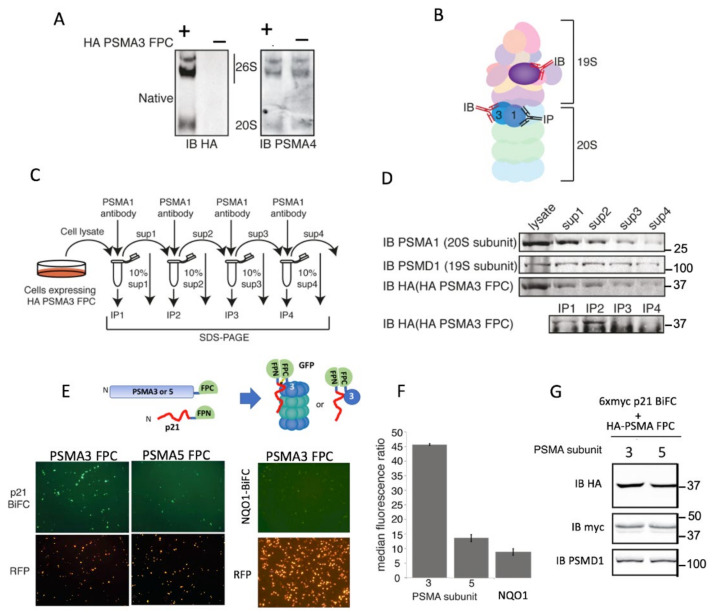
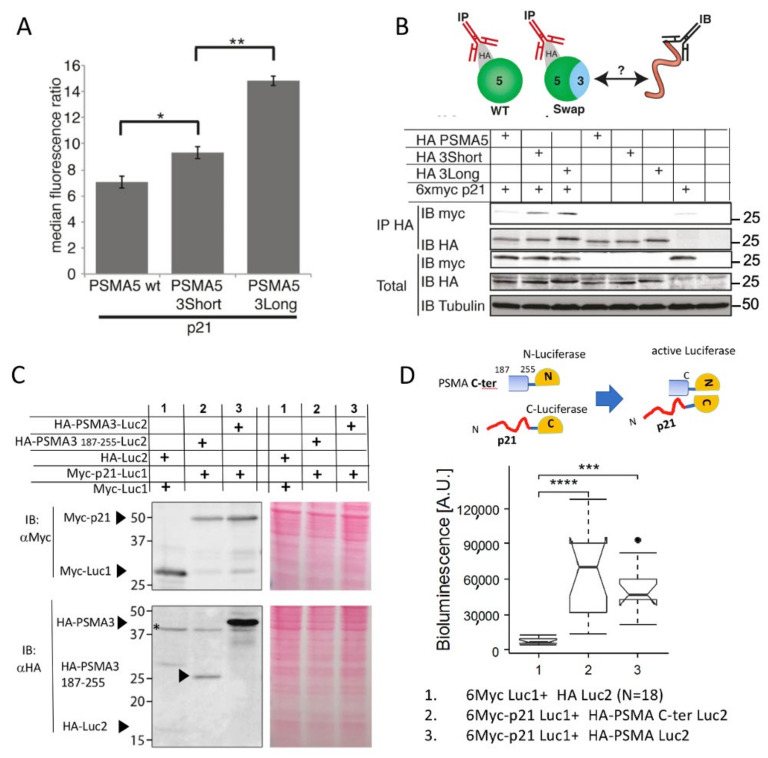
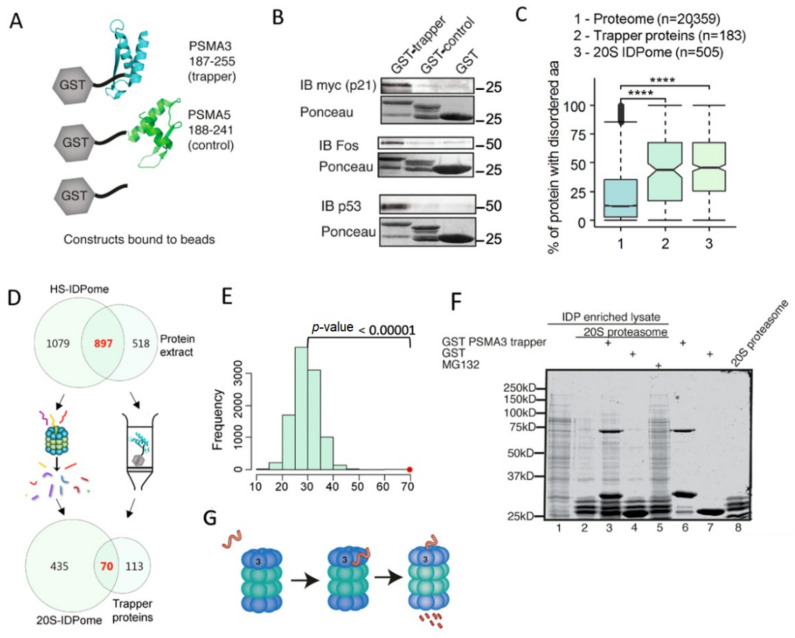
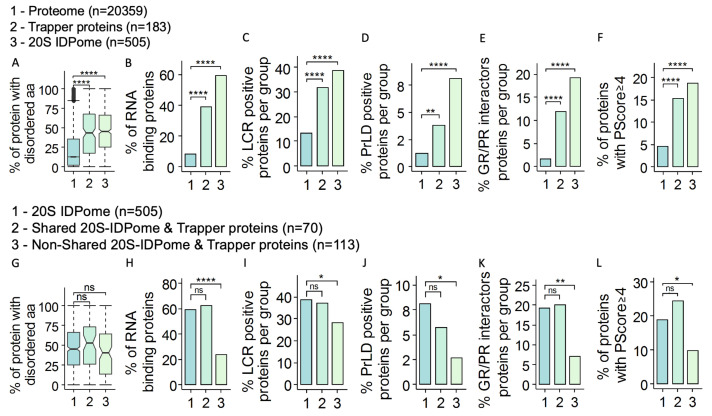
The degradation of intrinsically disordered proteins (IDPs) by a non-26S proteasome process does not require proteasomal targeting by polyubiquitin. However, whether and how IDPs are recognized by the non-26S proteasome, including the 20S complex, remains unknown. Analyses of protein interactome datasets revealed that the 20S proteasome subunit, PSMA3, preferentially interacts with many IDPs. In vivo and cell-free experiments revealed that the C-terminus of PSMA3, a 69-amino-acids-long fragment, is an IDP trapper. A recombinant trapper is sufficient to interact with many IDPs, and blocks IDP degradation in vitro by the 20S proteasome, possibly by competing with the native trapper. In addition, over a third of the PSMA3 trapper-binding proteins have previously been identified as 20S proteasome substrates and, based on published datasets, many of the trapper-binding proteins are associated with the intracellular proteasomes. The PSMA3-trapped IDPs that are proteasome substrates have the unique features previously recognized as characteristic 20S proteasome substrates in vitro. We propose a model whereby the PSMA3 C-terminal region traps a subset of IDPs to facilitate their proteasomal degradation.
Keywords: 20S proteasome; intrinsically disordered proteins; proteasomal degradation; proteostasis.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
