

Structural-Functional Relationship of the Ribonucleolytic Activity of aIF5A from Sulfolobus solfataricus
- PMID: 36291641
- PMCID: PMC9599864
- DOI: 10.3390/biom12101432
Structural-Functional Relationship of the Ribonucleolytic Activity of aIF5A from Sulfolobus solfataricus
Abstract
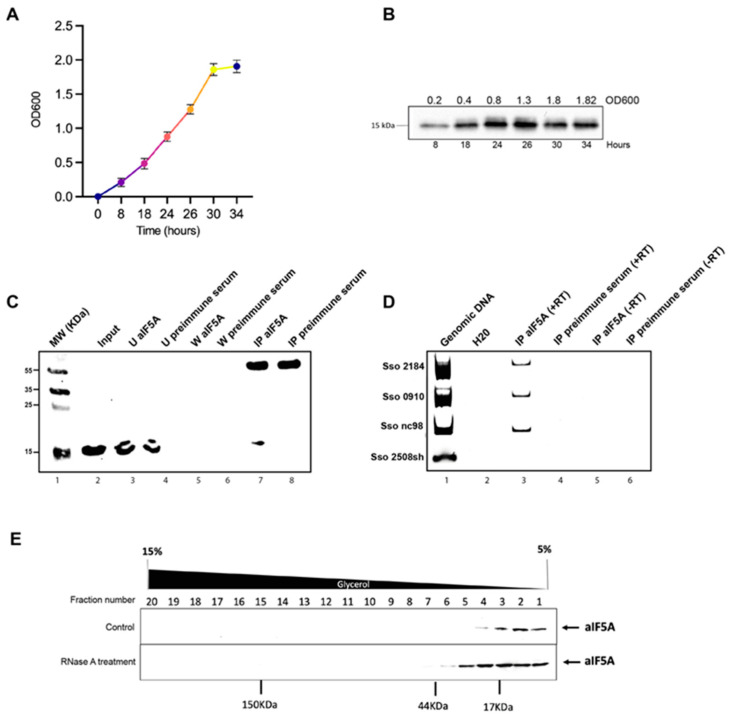
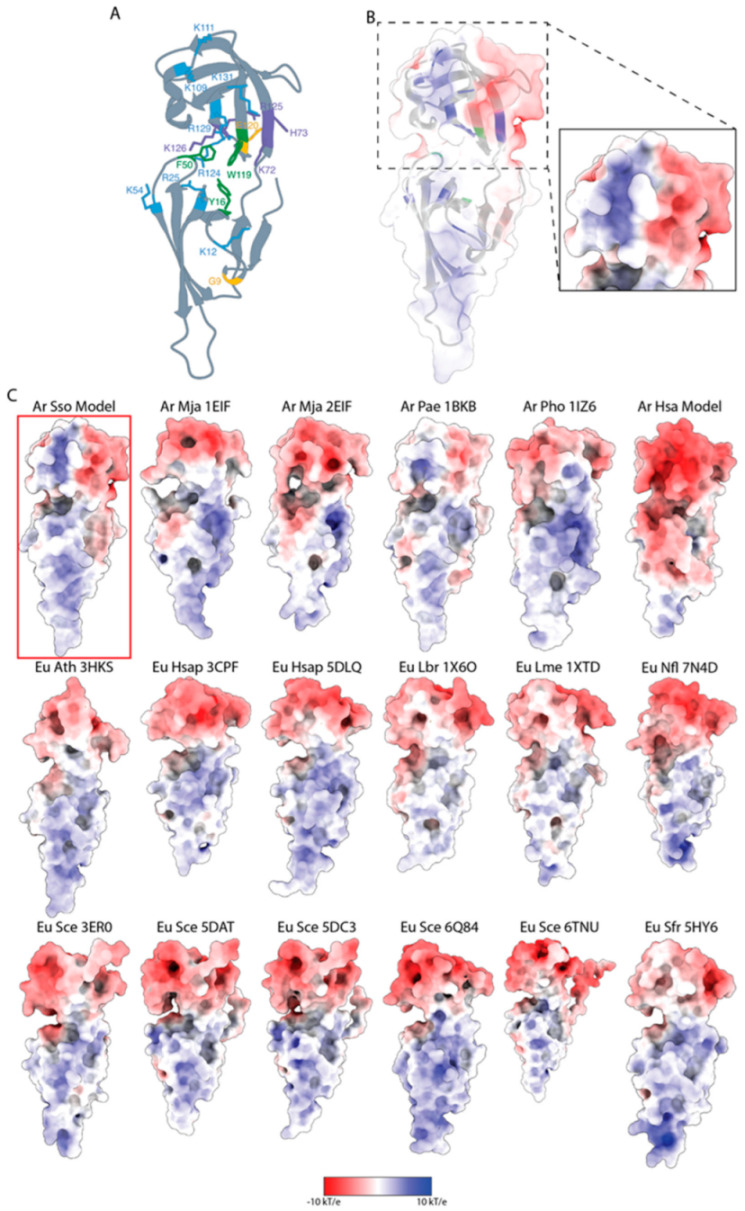
The translation factor IF5A is a highly conserved protein playing a well-recognized and well-characterized role in protein synthesis; nevertheless, some of its features as well as its abundance in the cell suggest that it may perform additional functions related to RNA metabolism. Here, we have undertaken a structural and functional characterization of aIF5A from the crenarchaeal Sulfolobus solfataricus model organism. We confirm the association of aIF5A with several RNA molecules in vivo and demonstrate that the protein is endowed with a ribonuclease activity which is specific for long and structured RNA. By means of biochemical and structural approaches we show that aIF5A can exist in both monomeric and dimeric conformations and the monomer formation is favored by the association with RNA. Finally, modelling of the three-dimensional structure of S. solfataricus aIF5A shows an extended positively charged surface which may explain its strong tendency to associate to RNA in vivo.
Keywords: Archaea; RNA metabolism; SAXS; Sulfolobus solfataricus; aIF5A; ribonuclease.
Conflict of interest statement
The authors declare no conflict of interest.
Figures



References
-
- Park J.H., Dias C.A.O., Lee S.B., Valentini S.R., Sokabe M., Fraser C.S., Park M.H. Production of Active Recombinant EIF5A: Reconstitution in E.Coli of Eukaryotic Hypusine Modification of EIF5A by Its Coexpression with Modifying Enzymes. Protein Eng. Des. Sel. 2011;24:301–309. doi: 10.1093/protein/gzq110. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
