

Chromatin as a sensor of metabolic changes during early development
- PMID: 36299478
- PMCID: PMC9588933
- DOI: 10.3389/fcell.2022.1014498
Chromatin as a sensor of metabolic changes during early development
Abstract
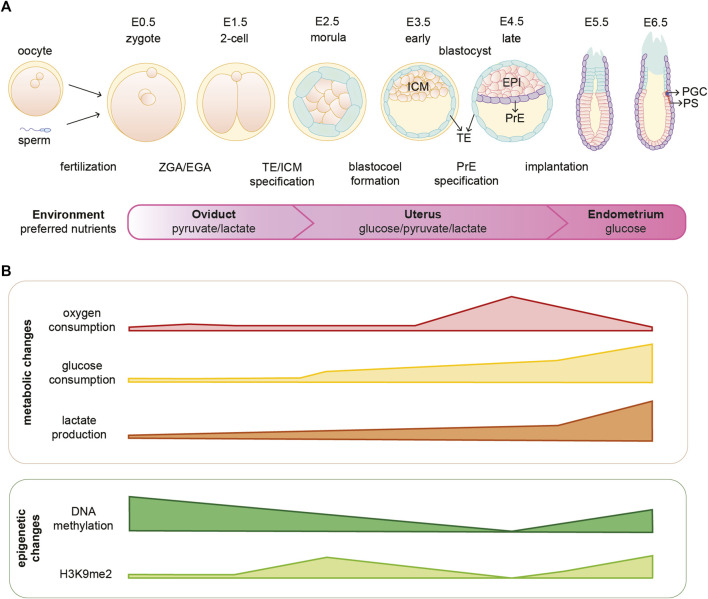
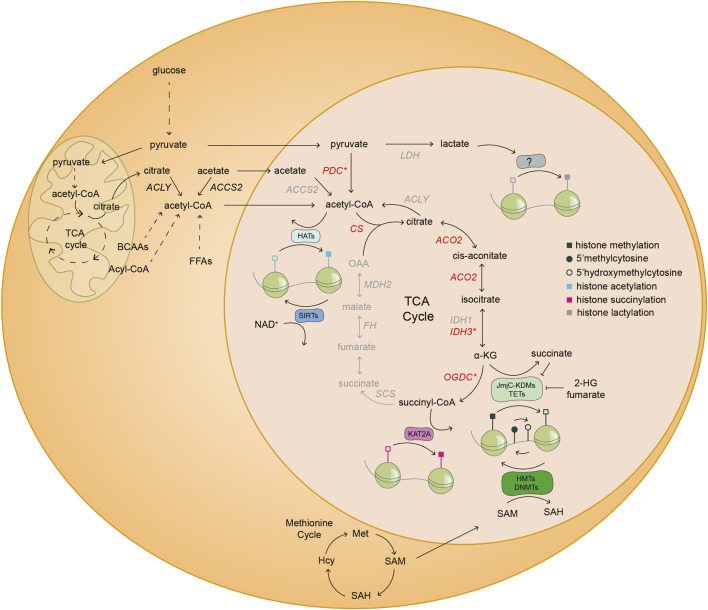
Cellular metabolism is a complex network of biochemical reactions fueling development with energy and biomass; however, it can also shape the cellular epigenome. Indeed, some intermediates of metabolic reactions exert a non-canonical function by acting as co-factors, substrates or inhibitors of chromatin modifying enzymes. Therefore, fluctuating availability of such molecules has the potential to regulate the epigenetic landscape. Thanks to this functional coupling, chromatin can act as a sensor of metabolic changes and thus impact cell fate. Growing evidence suggest that both metabolic and epigenetic reprogramming are crucial for ensuring a successful embryo development from the zygote until gastrulation. In this review, we provide an overview of the complex relationship between metabolism and epigenetics in regulating the early stages of mammalian embryo development. We report on recent breakthroughs in uncovering the non-canonical functions of metabolism especially when re-localized to the nucleus. In addition, we identify the challenges and outline future perspectives to advance the novel field of epi-metabolomics especially in the context of early development.
Keywords: cell fate; chromatin; early embryonic development; epi-metabolomics; epigenetics; metabolism; nuclear metabolism.
Copyright © 2022 Pladevall-Morera and Zylicz.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
Similar articles
-
Metabolism-epigenetic interactions on in vitro produced embryos.Reprod Fertil Dev. 2022 Dec;35(2):84-97. doi: 10.1071/RD22203. Reprod Fertil Dev. 2022. PMID: 36592974 Review.
-
Metabolic regulation of the plant epigenome.Plant J. 2023 Jun;114(5):1001-1013. doi: 10.1111/tpj.16122. Epub 2023 Feb 10. Plant J. 2023. PMID: 36705504 Review.
-
The Bidirectional Relationship Between Cancer Epigenetics and Metabolism.Annu Rev Cancer Biol. 2021 Mar;5(1):235-257. doi: 10.1146/annurev-cancerbio-070820-035832. Epub 2020 Nov 30. Annu Rev Cancer Biol. 2021. PMID: 34109280 Free PMC article.
-
Crosstalk between metabolic reprogramming and epigenetics in cancer: updates on mechanisms and therapeutic opportunities.Cancer Commun (Lond). 2022 Nov;42(11):1049-1082. doi: 10.1002/cac2.12374. Epub 2022 Oct 20. Cancer Commun (Lond). 2022. PMID: 36266736 Free PMC article. Review.
-
[The inheritance and reprogramming of chromatin structure in early animal embryos].Yi Chuan. 2018 Nov 20;40(11):977-987. doi: 10.16288/j.yczz.18-189. Yi Chuan. 2018. PMID: 30465530 Review. Chinese.
Cited by
-
Glucose-Induced Developmental Dynamics: Understanding Male Prevalence in Early Mouse Embryo Stages.Reprod Med Biol. 2025 Jul 5;24(1):e12667. doi: 10.1002/rmb2.12667. eCollection 2025 Jan-Dec. Reprod Med Biol. 2025. PMID: 40620426 Free PMC article.
-
α-Ketoglutarate promotes trophectoderm induction and maturation from naive human embryonic stem cells.Nat Cell Biol. 2025 May;27(5):749-761. doi: 10.1038/s41556-025-01658-1. Epub 2025 Apr 23. Nat Cell Biol. 2025. PMID: 40269259 Free PMC article.
-
Molecular mechanisms of cellular metabolic homeostasis in stem cells.Int J Oral Sci. 2023 Dec 1;15(1):52. doi: 10.1038/s41368-023-00262-z. Int J Oral Sci. 2023. PMID: 38040705 Free PMC article. Review.
References
Publication types
LinkOut - more resources
Full Text Sources