

Commensal microbiota from patients with inflammatory bowel disease produce genotoxic metabolites
- PMID: 36302024
- PMCID: PMC9993714
- DOI: 10.1126/science.abm3233
Commensal microbiota from patients with inflammatory bowel disease produce genotoxic metabolites
Abstract
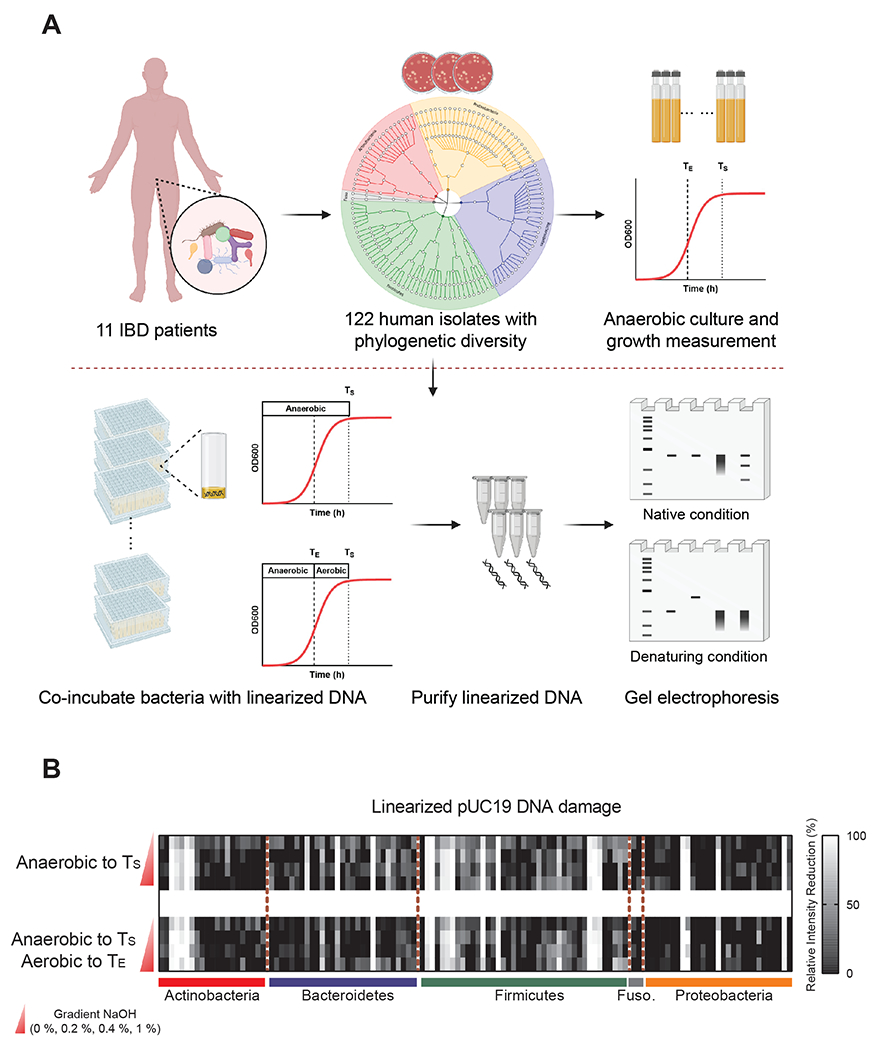
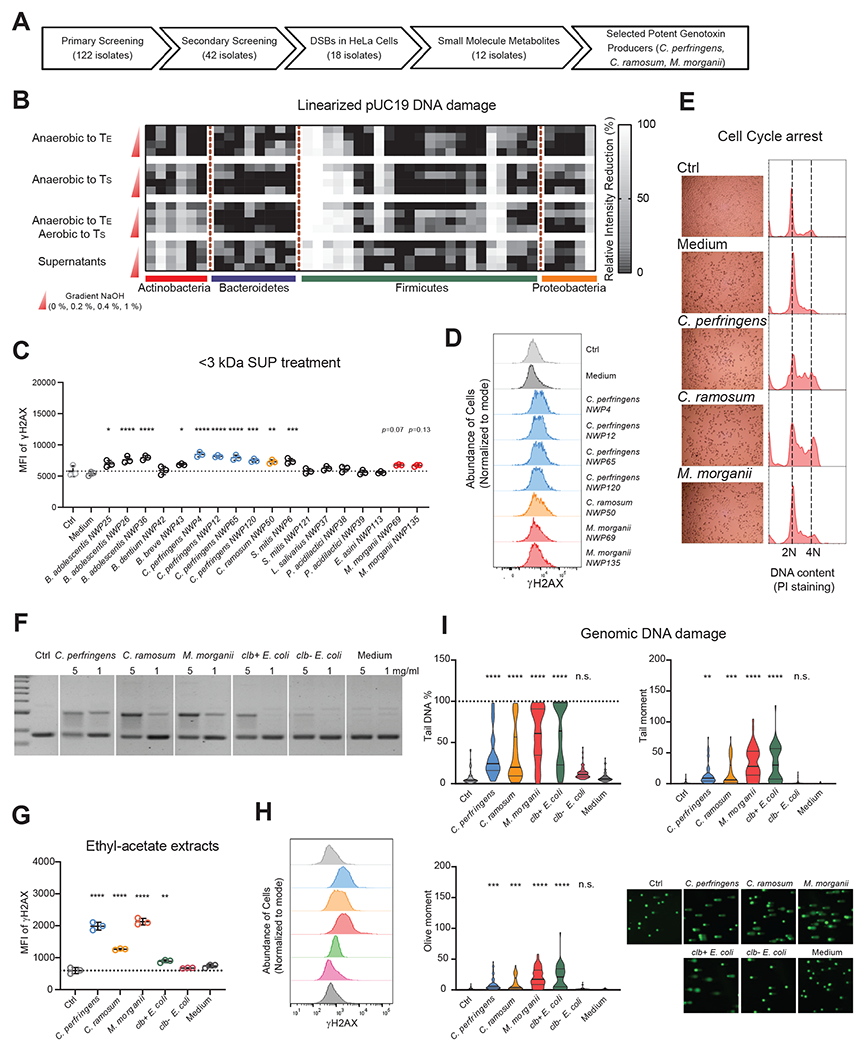
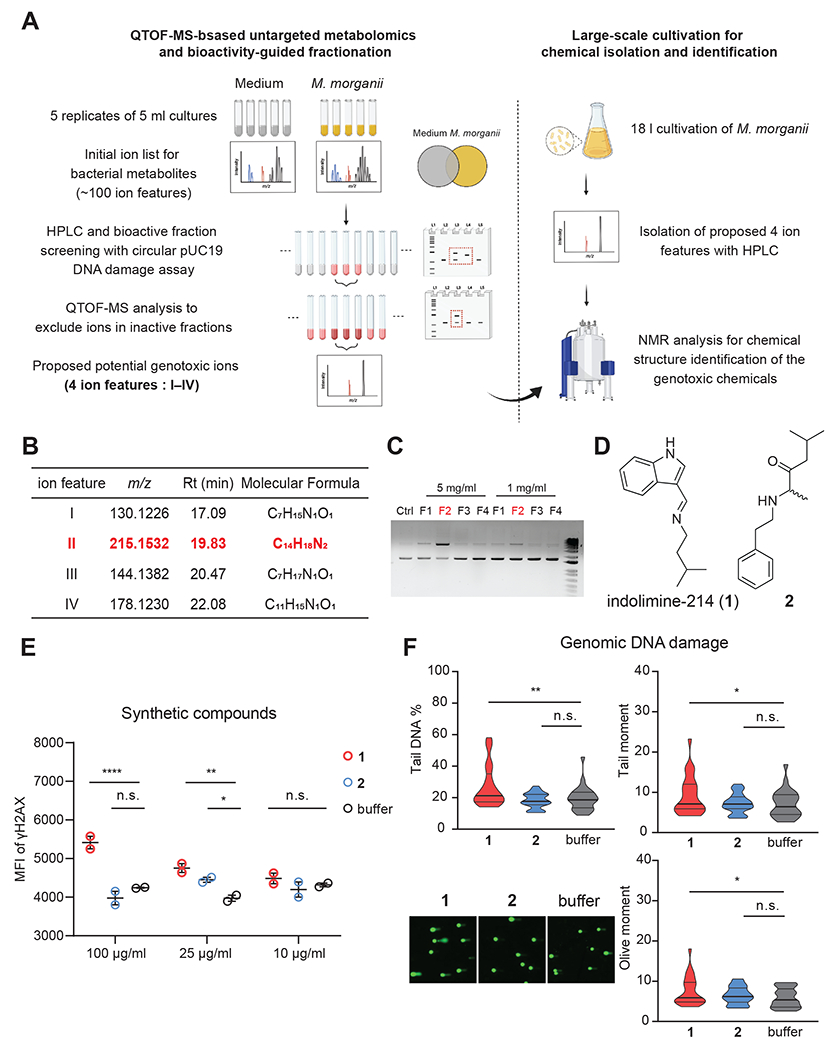
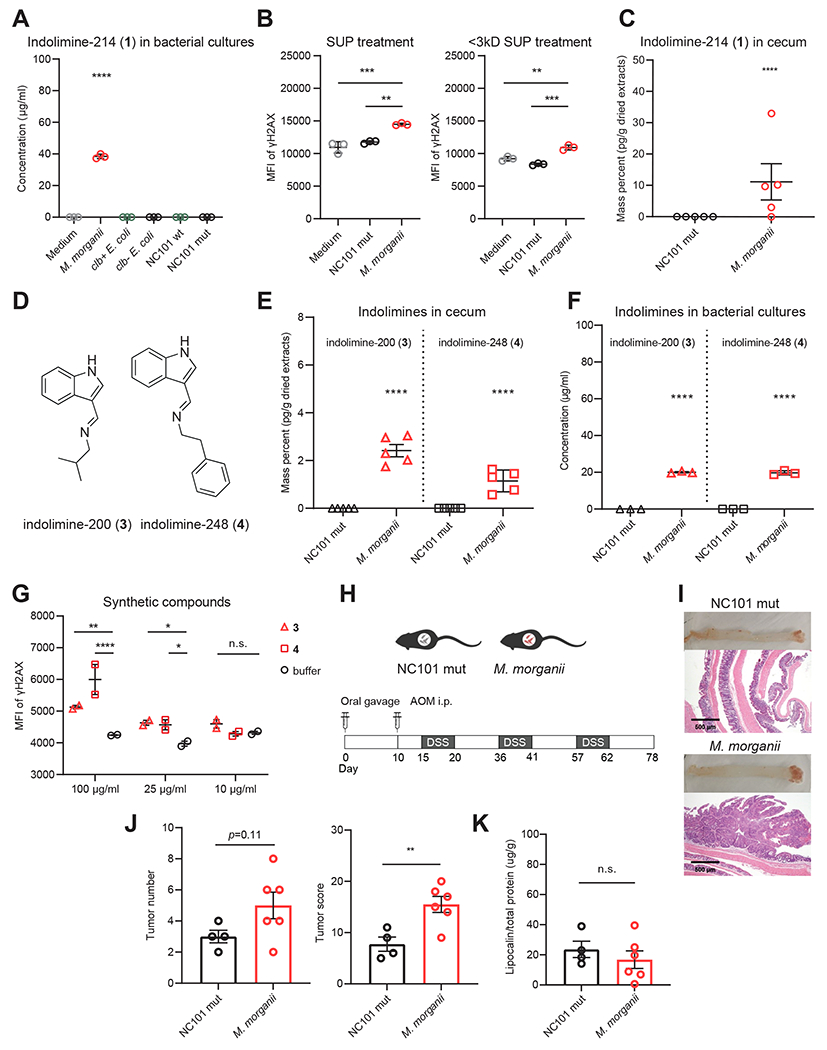
Microbiota-derived metabolites that elicit DNA damage can contribute to colorectal cancer (CRC). However, the full spectrum of genotoxic chemicals produced by indigenous gut microbes remains to be defined. We established a pipeline to systematically evaluate the genotoxicity of an extensive collection of gut commensals from inflammatory bowel disease patients. We identified isolates from divergent phylogenies whose metabolites caused DNA damage and discovered a distinctive family of genotoxins-termed the indolimines-produced by the CRC-associated species Morganella morganii. A non-indolimine-producing M. morganii mutant lacked genotoxicity and failed to exacerbate colon tumorigenesis in mice. These studies reveal the existence of a previously unexplored universe of genotoxic small molecules from the microbiome that may affect host biology in homeostasis and disease.
Figures
Comment in
-
Microbial metabolites damage DNA.Science. 2022 Oct 28;378(6618):358-359. doi: 10.1126/science.ade6952. Epub 2022 Oct 27. Science. 2022. PMID: 36302018
-
Gut microbes produce novel genotoxic metabolites.Nat Rev Cancer. 2023 Jan;23(1):3. doi: 10.1038/s41568-022-00538-2. Nat Rev Cancer. 2023. PMID: 36450856 No abstract available.
References
-
- Keum N, Giovannucci E, Global burden of colorectal cancer: emerging trends, risk factors and prevention strategies. Nat Rev Gastroenterol Hepatol 16, 713–732 (2019). - PubMed
-
- Tilg H, Adolph TE, Gerner RR, Moschen AR, The Intestinal Microbiota in Colorectal Cancer. Cancer Cell 33, 954–964 (2018). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
