

Regressive evolution of an effector following a host jump in the Irish potato famine pathogen lineage
- PMID: 36302035
- PMCID: PMC9642902
- DOI: 10.1371/journal.ppat.1010918
Regressive evolution of an effector following a host jump in the Irish potato famine pathogen lineage
Abstract
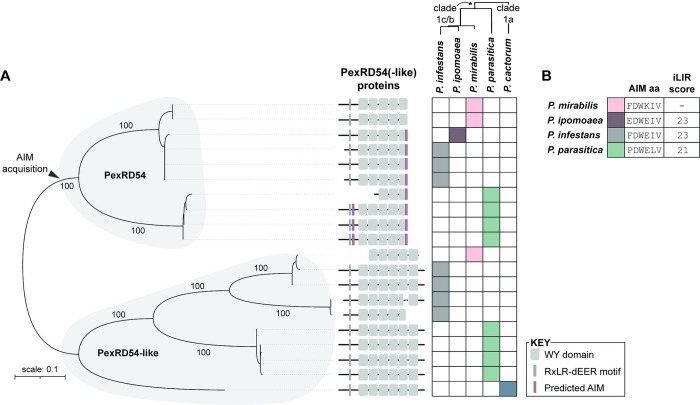
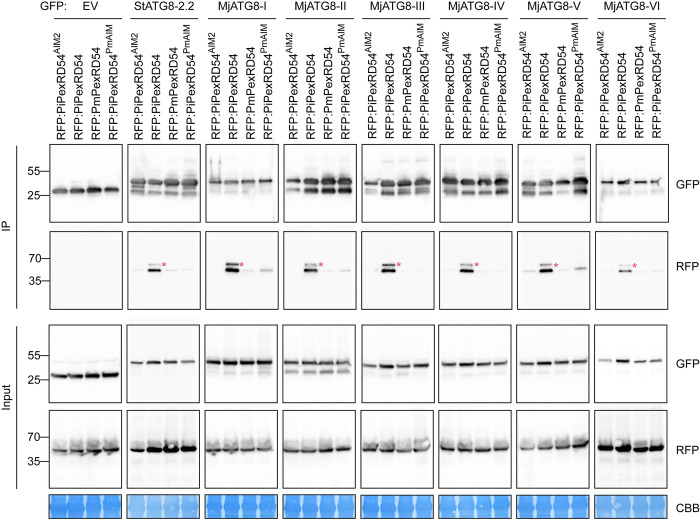
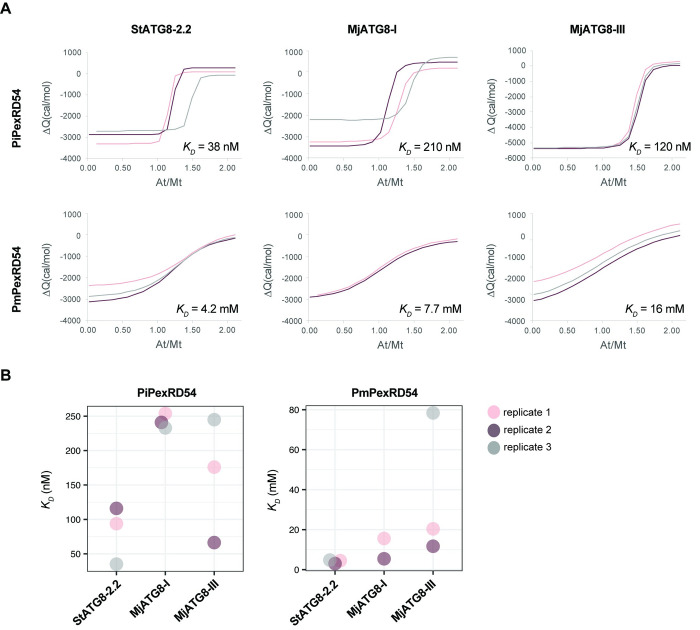
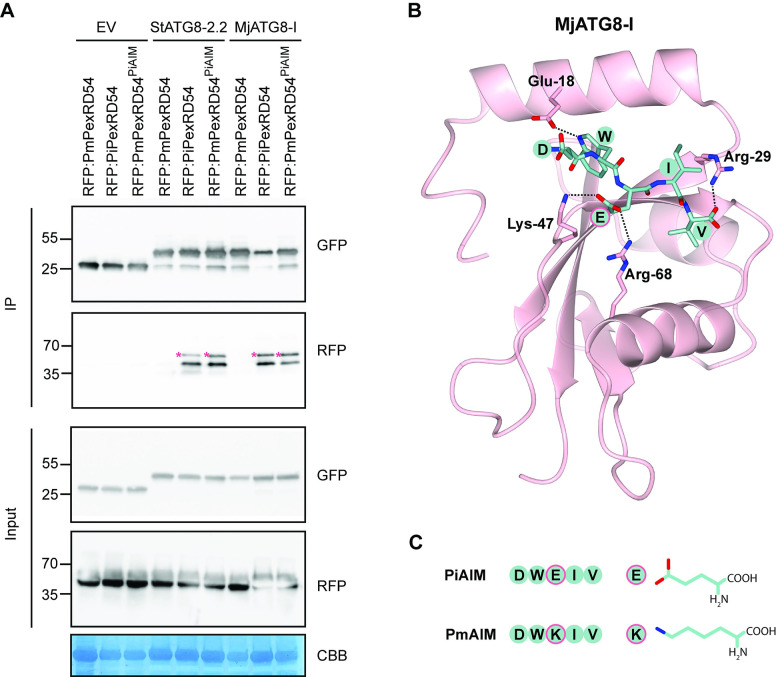
In order to infect a new host species, the pathogen must evolve to enhance infection and transmission in the novel environment. Although we often think of evolution as a process of accumulation, it is also a process of loss. Here, we document an example of regressive evolution of an effector activity in the Irish potato famine pathogen (Phytophthora infestans) lineage, providing evidence that a key sequence motif in the effector PexRD54 has degenerated following a host jump. We began by looking at PexRD54 and PexRD54-like sequences from across Phytophthora species. We found that PexRD54 emerged in the common ancestor of Phytophthora clade 1b and 1c species, and further sequence analysis showed that a key functional motif, the C-terminal ATG8-interacting motif (AIM), was also acquired at this point in the lineage. A closer analysis showed that the P. mirabilis PexRD54 (PmPexRD54) AIM is atypical, the otherwise-conserved central residue mutated from a glutamate to a lysine. We aimed to determine whether this PmPexRD54 AIM polymorphism represented an adaptation to the Mirabilis jalapa host environment. We began by characterizing the M. jalapa ATG8 family, finding that they have a unique evolutionary history compared to previously characterized ATG8s. Then, using co-immunoprecipitation and isothermal titration calorimetry assays, we showed that both full-length PmPexRD54 and the PmPexRD54 AIM peptide bind weakly to the M. jalapa ATG8s. Through a combination of binding assays and structural modelling, we showed that the identity of the residue at the position of the PmPexRD54 AIM polymorphism can underpin high-affinity binding to plant ATG8s. Finally, we conclude that the functionality of the PexRD54 AIM was lost in the P. mirabilis lineage, perhaps owing to as-yet-unknown selection pressure on this effector in the new host environment.
Copyright: © 2022 Zess et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures


References
-
- Parker IM, Gilbert GS. The Evolutionary Ecology of Novel Plant-Pathogen Interactions. Annu Rev Ecol Evol Syst. 2004. Dec 15;35(1):675–700.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
