

Towards a unified theory of plant photosynthesis and hydraulics
- PMID: 36303010
- PMCID: PMC9663302
- DOI: 10.1038/s41477-022-01244-5
Towards a unified theory of plant photosynthesis and hydraulics
Abstract
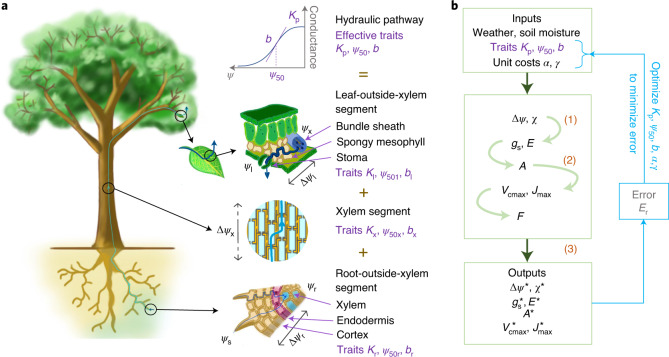
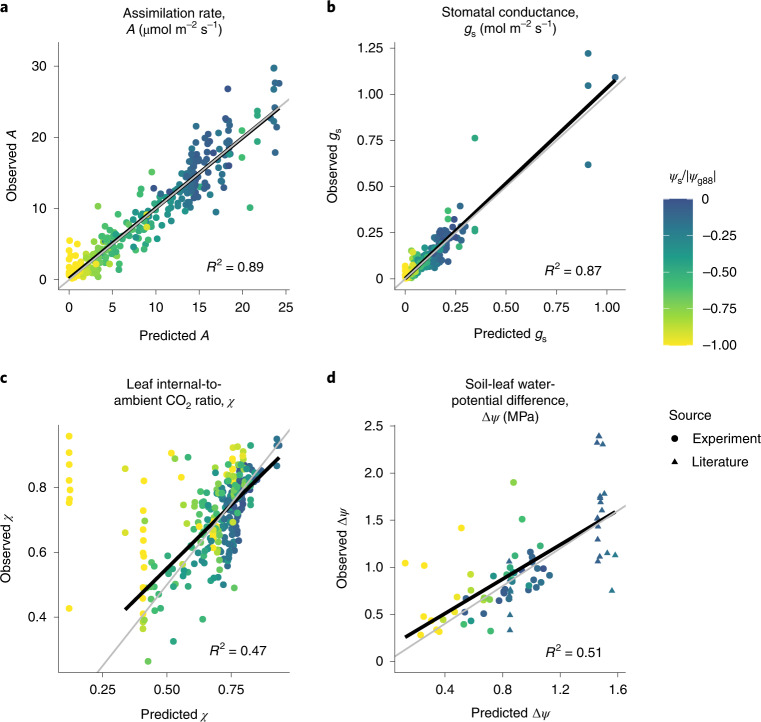
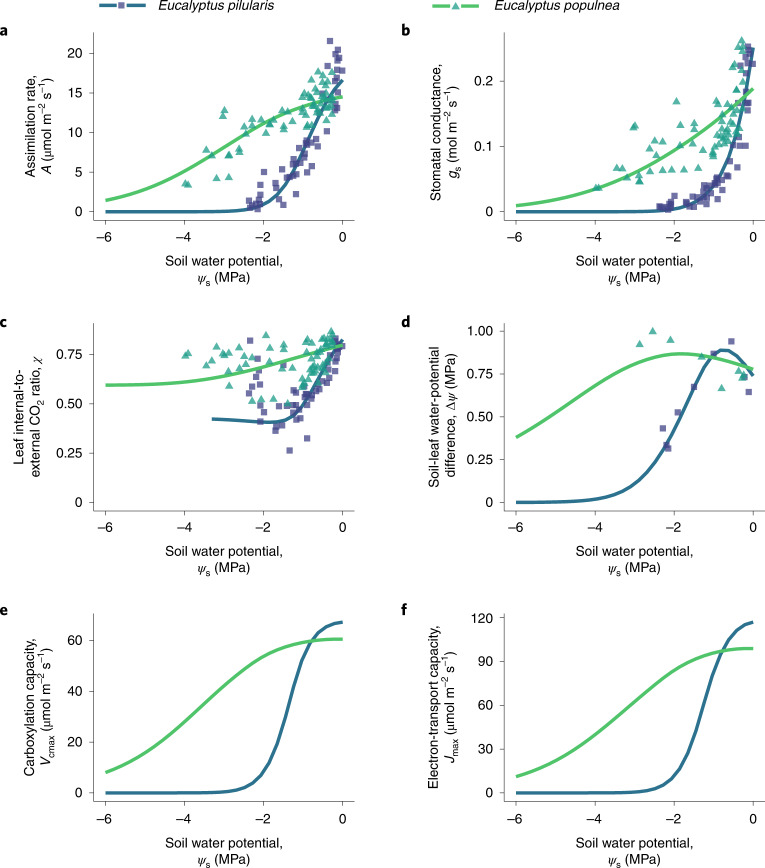
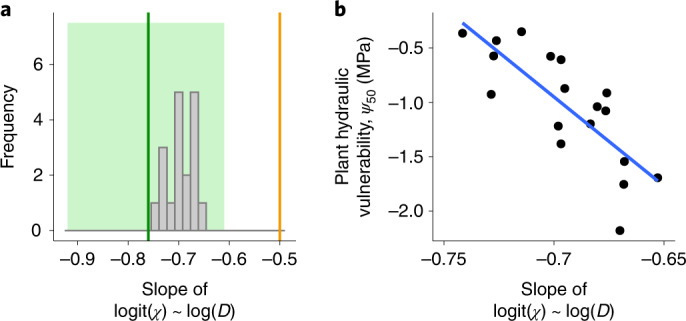
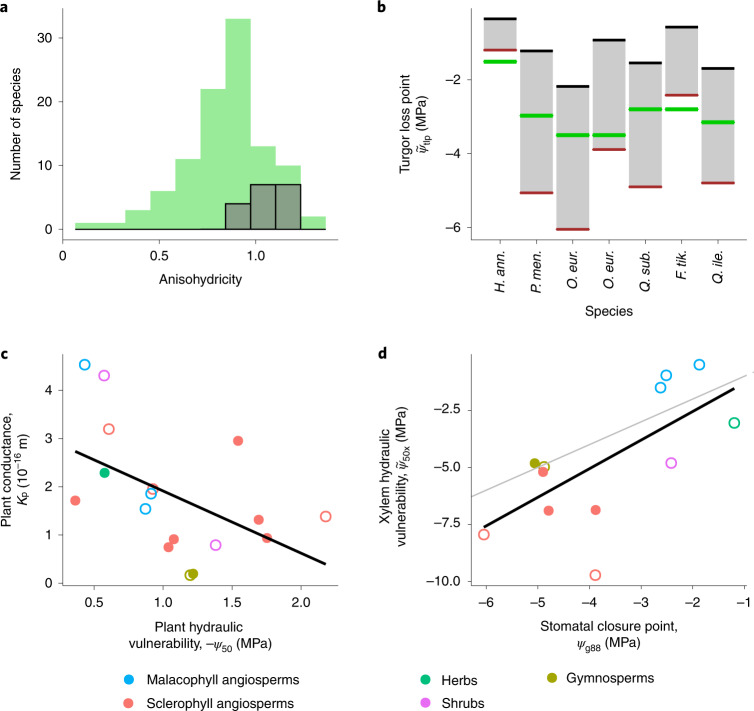
The global carbon and water cycles are governed by the coupling of CO2 and water vapour exchanges through the leaves of terrestrial plants, controlled by plant adaptations to balance carbon gains and hydraulic risks. We introduce a trait-based optimality theory that unifies the treatment of stomatal responses and biochemical acclimation of plants to environments changing on multiple timescales. Tested with experimental data from 18 species, our model successfully predicts the simultaneous decline in carbon assimilation rate, stomatal conductance and photosynthetic capacity during progressive soil drought. It also correctly predicts the dependencies of gas exchange on atmospheric vapour pressure deficit, temperature and CO2. Model predictions are also consistent with widely observed empirical patterns, such as the distribution of hydraulic strategies. Our unified theory opens new avenues for reliably modelling the interactive effects of drying soil and rising atmospheric CO2 on global photosynthesis and transpiration.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Raschke K, Monteith JL, Weatherley PE. How stomata resolve the dilemma of opposing priorities. Phil. Trans. R. Soc. Lond. B. 1976;273:551–560. doi: 10.1098/rstb.1976.0031. - DOI
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
