

High-throughput automated methods for classical and operant conditioning of Drosophila larvae
- PMID: 36305588
- PMCID: PMC9678368
- DOI: 10.7554/eLife.70015
High-throughput automated methods for classical and operant conditioning of Drosophila larvae
Abstract
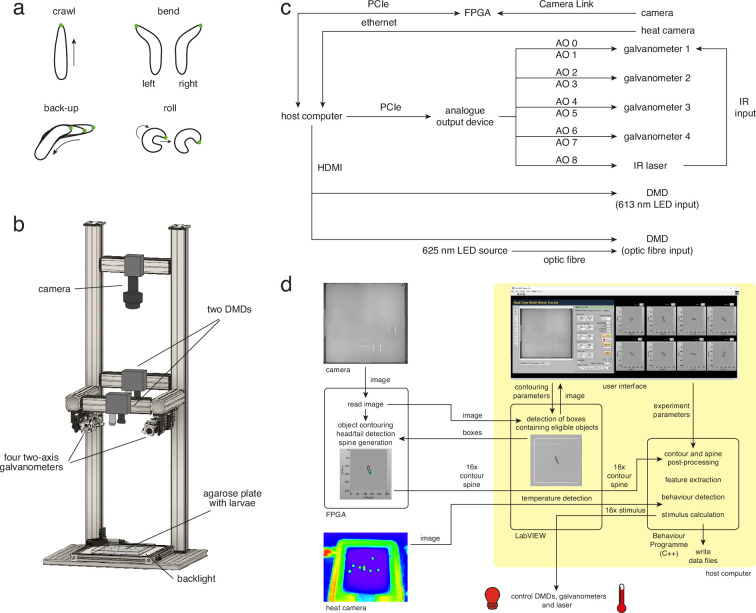
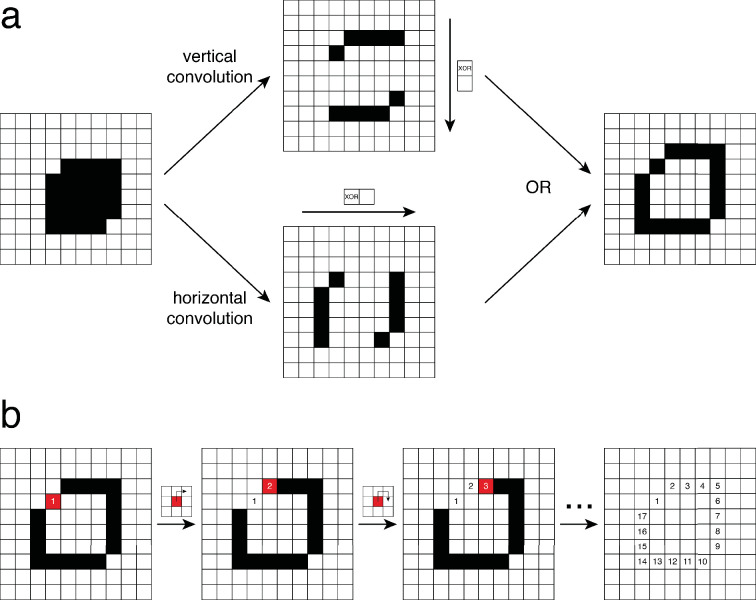
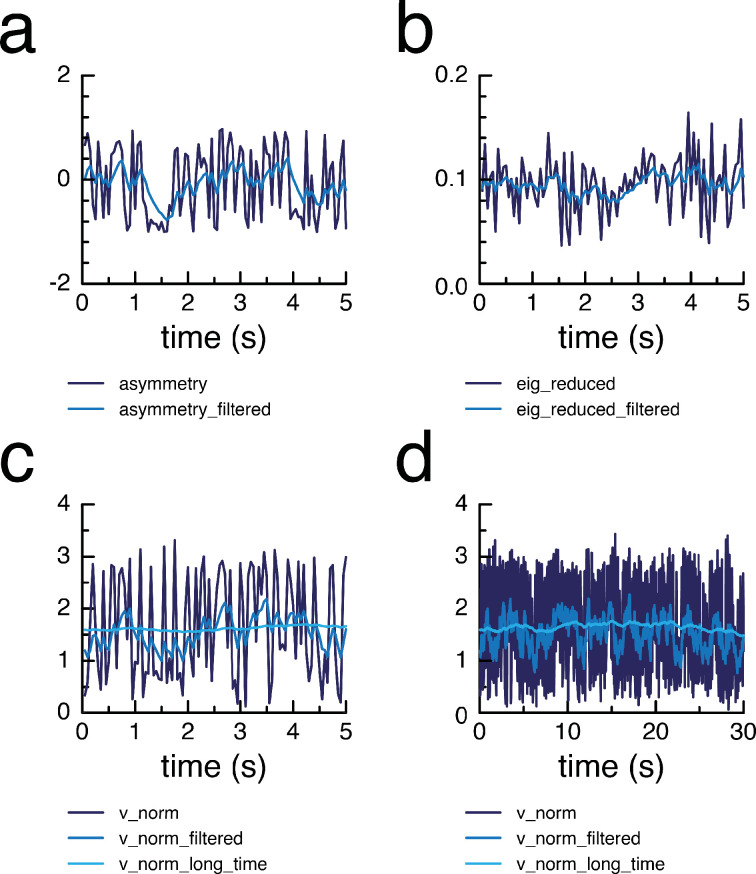
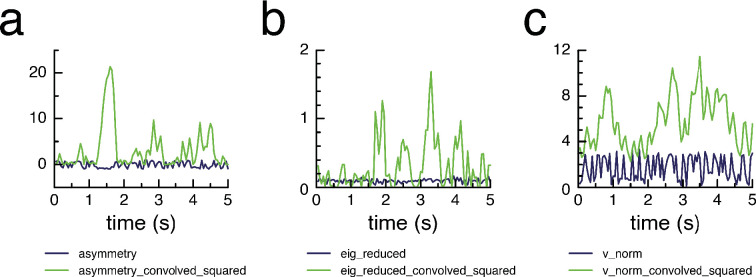
Learning which stimuli (classical conditioning) or which actions (operant conditioning) predict rewards or punishments can improve chances of survival. However, the circuit mechanisms that underlie distinct types of associative learning are still not fully understood. Automated, high-throughput paradigms for studying different types of associative learning, combined with manipulation of specific neurons in freely behaving animals, can help advance this field. The Drosophila melanogaster larva is a tractable model system for studying the circuit basis of behaviour, but many forms of associative learning have not yet been demonstrated in this animal. Here, we developed a high-throughput (i.e. multi-larva) training system that combines real-time behaviour detection of freely moving larvae with targeted opto- and thermogenetic stimulation of tracked animals. Both stimuli are controlled in either open- or closed-loop, and delivered with high temporal and spatial precision. Using this tracker, we show for the first time that Drosophila larvae can perform classical conditioning with no overlap between sensory stimuli (i.e. trace conditioning). We also demonstrate that larvae are capable of operant conditioning by inducing a bend direction preference through optogenetic activation of reward-encoding serotonergic neurons. Our results extend the known associative learning capacities of Drosophila larvae. Our automated training rig will facilitate the study of many different forms of associative learning and the identification of the neural circuits that underpin them.
Keywords: D. melanogaster; Drosophila melanogaster larva; behavior detection; closed-loop tracking; computational biology; computer vision; neuroscience; operant conditioning; stimulation; systems biology; trace conditioning.
© 2022, Croteau-Chonka, Clayton et al.
Conflict of interest statement
EC, MC, LV, SH, BJ, LN, MW, JM, MZ, KK No competing interests declared
Figures













Similar articles
-
Drosophila larvae form appetitive and aversive associative memory in response to thermal conditioning.PLoS One. 2024 Sep 24;19(9):e0303955. doi: 10.1371/journal.pone.0303955. eCollection 2024. PLoS One. 2024. PMID: 39316589 Free PMC article.
-
Switch-like and persistent memory formation in individual Drosophila larvae.Elife. 2021 Oct 12;10:e70317. doi: 10.7554/eLife.70317. Elife. 2021. PMID: 34636720 Free PMC article.
-
Feeding behavior of Aplysia: a model system for comparing cellular mechanisms of classical and operant conditioning.Learn Mem. 2006 Nov-Dec;13(6):669-80. doi: 10.1101/lm.339206. Learn Mem. 2006. PMID: 17142299 Review.
-
The operant and the classical in conditioned orientation of Drosophila melanogaster at the flight simulator.Learn Mem. 2000 Mar-Apr;7(2):104-15. doi: 10.1101/lm.7.2.104. Learn Mem. 2000. PMID: 10753977 Free PMC article.
-
Functional basis of associative learning and its relationships with long-term potentiation evoked in the involved neural circuits: Lessons from studies in behaving mammals.Neurobiol Learn Mem. 2015 Oct;124:3-18. doi: 10.1016/j.nlm.2015.04.006. Epub 2015 Apr 25. Neurobiol Learn Mem. 2015. PMID: 25916668 Review.
Cited by
-
Neuronal circuit mechanisms of competitive interaction between action-based and coincidence learning.Sci Adv. 2024 Dec 6;10(49):eadq3016. doi: 10.1126/sciadv.adq3016. Epub 2024 Dec 6. Sci Adv. 2024. PMID: 39642217 Free PMC article.
-
Drosophila larvae form appetitive and aversive associative memory in response to thermal conditioning.PLoS One. 2024 Sep 24;19(9):e0303955. doi: 10.1371/journal.pone.0303955. eCollection 2024. PLoS One. 2024. PMID: 39316589 Free PMC article.
-
Statistical signature of subtle behavioral changes in large-scale assays.PLoS Comput Biol. 2025 Apr 21;21(4):e1012990. doi: 10.1371/journal.pcbi.1012990. eCollection 2025 Apr. PLoS Comput Biol. 2025. PMID: 40258220 Free PMC article.
-
High-resolution analysis of individual Drosophila melanogaster larvae uncovers individual variability in locomotion and its neurogenetic modulation.Open Biol. 2023 Apr;13(4):220308. doi: 10.1098/rsob.220308. Epub 2023 Apr 19. Open Biol. 2023. PMID: 37072034 Free PMC article.
-
Future avenues in Drosophila mushroom body research.Learn Mem. 2024 Jun 11;31(5):a053863. doi: 10.1101/lm.053863.123. Print 2024 May. Learn Mem. 2024. PMID: 38862172 Free PMC article. Review.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
