

Testis-specific serine kinase 3 is required for sperm morphogenesis and male fertility
- PMID: 36306217
- PMCID: PMC10267670
- DOI: 10.1111/andr.13314
Testis-specific serine kinase 3 is required for sperm morphogenesis and male fertility
Abstract
Background: The importance of phosphorylation in sperm during spermatogenesis has not been pursued extensively. Testis-specific serine kinase 3 (Tssk3) is a conserved gene, but TSSK3 kinase functions and phosphorylation substrates of TSSK3 are not known.
Objective: The goals of our studies were to understand the mechanism of action of TSSK3.
Materials and methods: We analyzed the localization of TSSK3 in sperm, used CRISPR/Cas9 to generate Tssk3 knockout (KO) mice in which nearly all of the Tssk3 open reading frame was deleted (ensuring it is a null mutation), analyzed the fertility of Tssk3 KO mice by breeding mice for 4 months, and conducted phosphoproteomics analysis of male testicular germ cells.
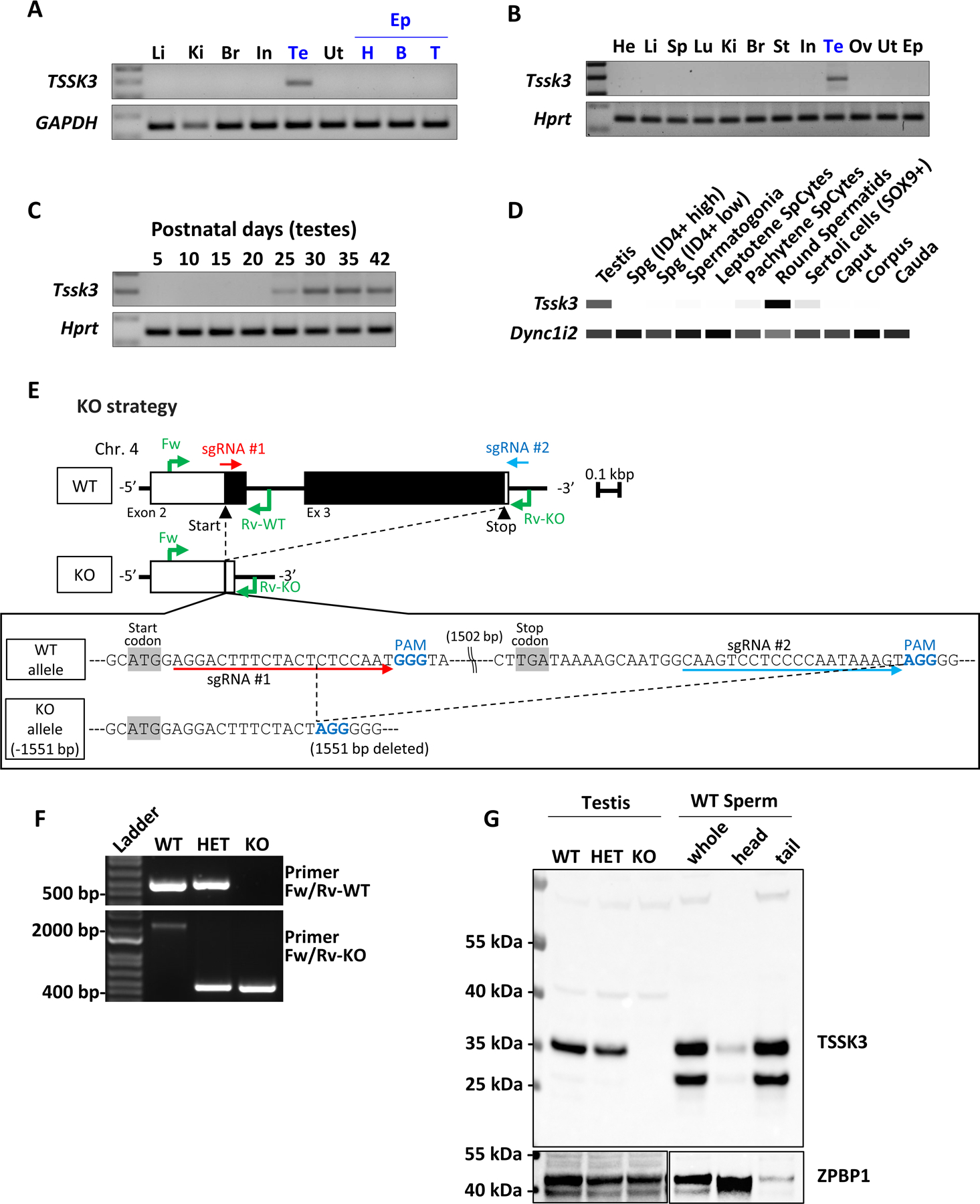
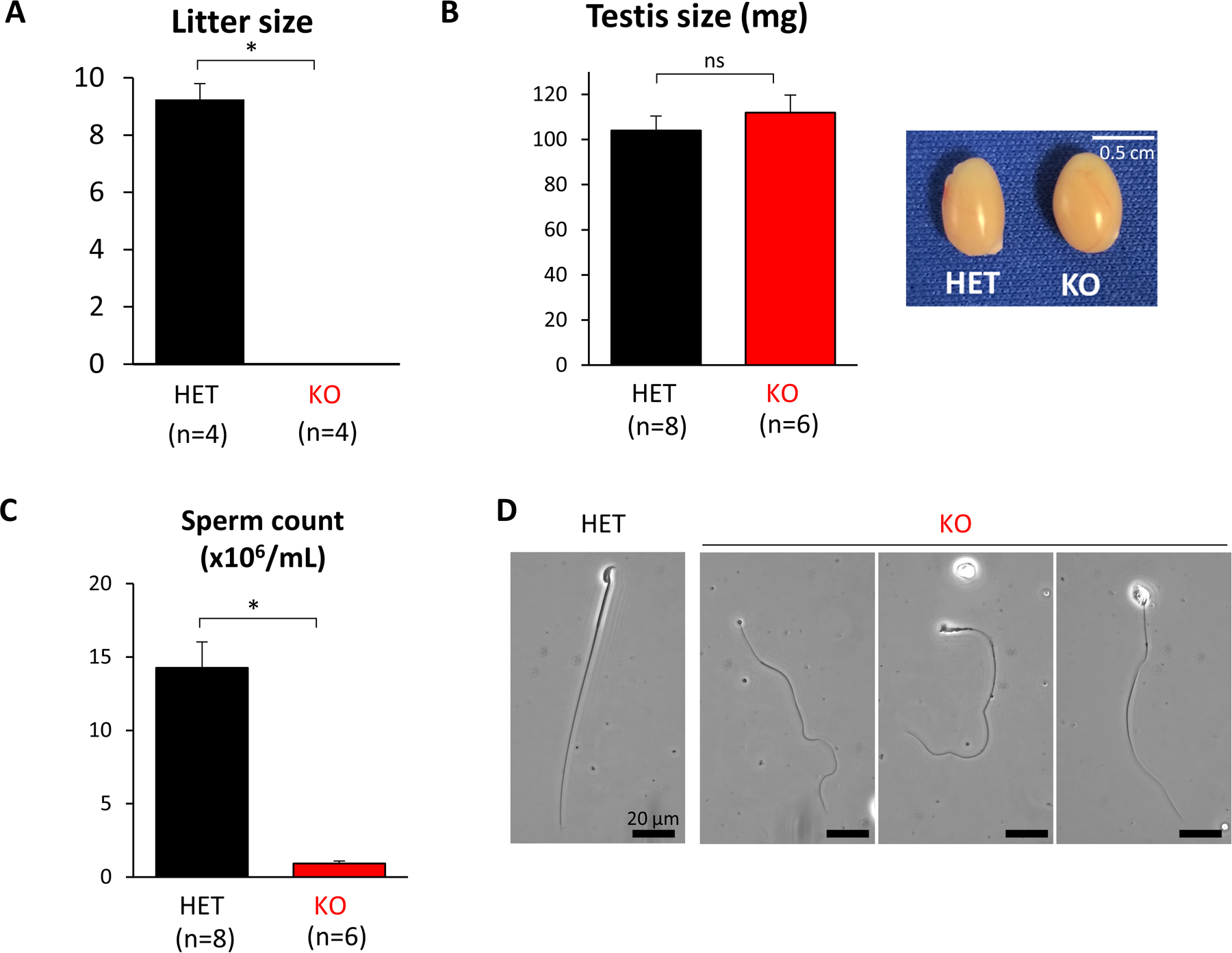
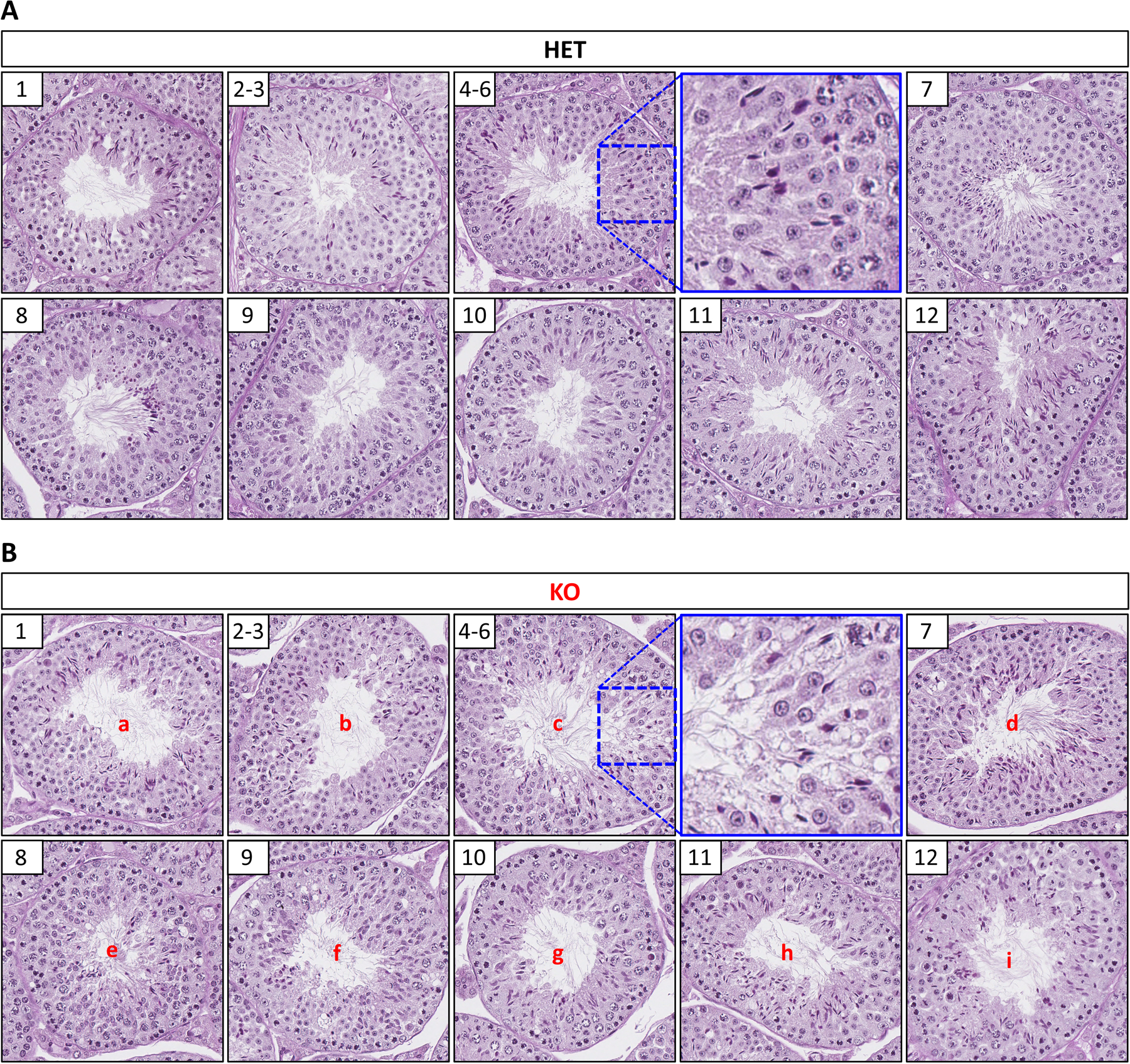
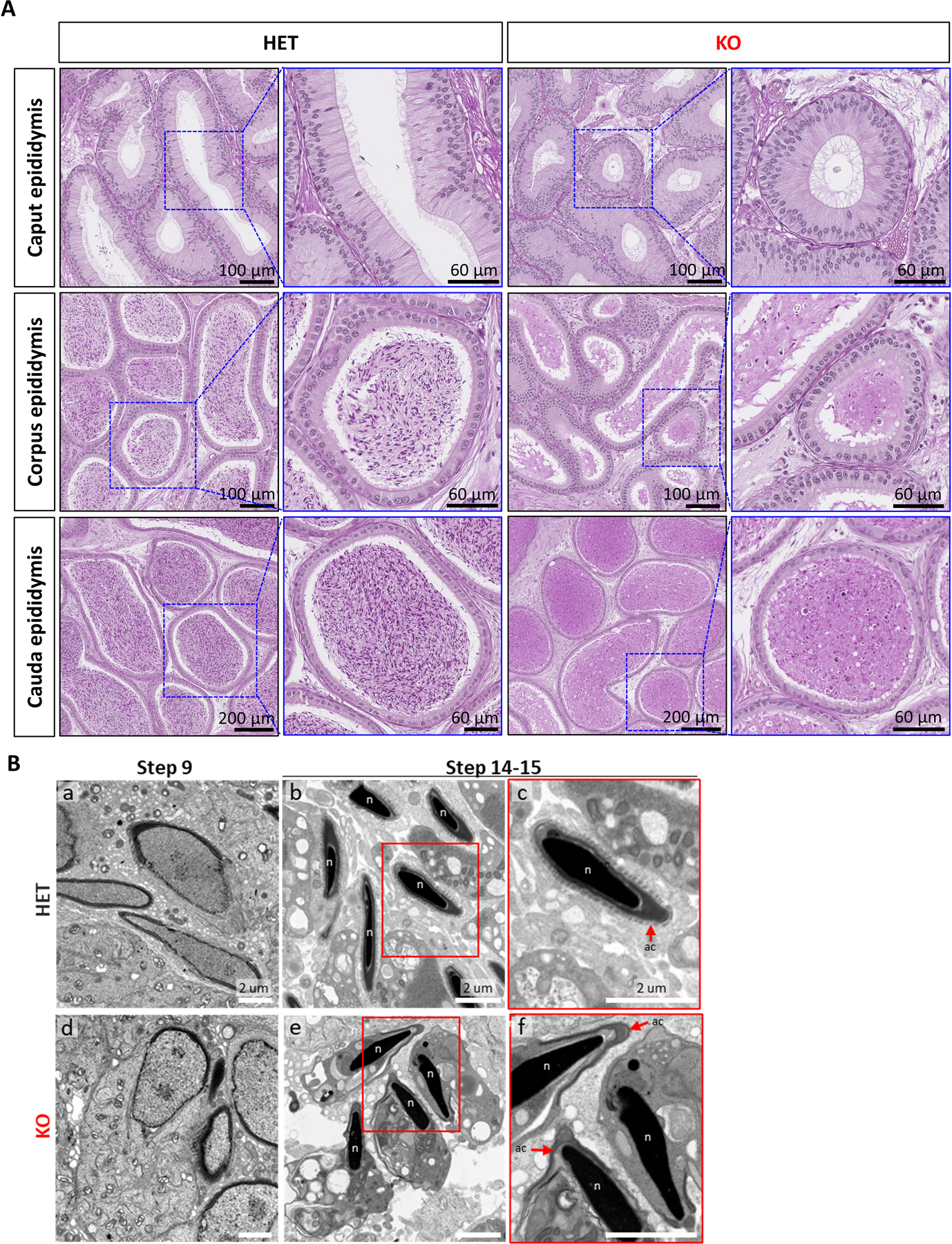
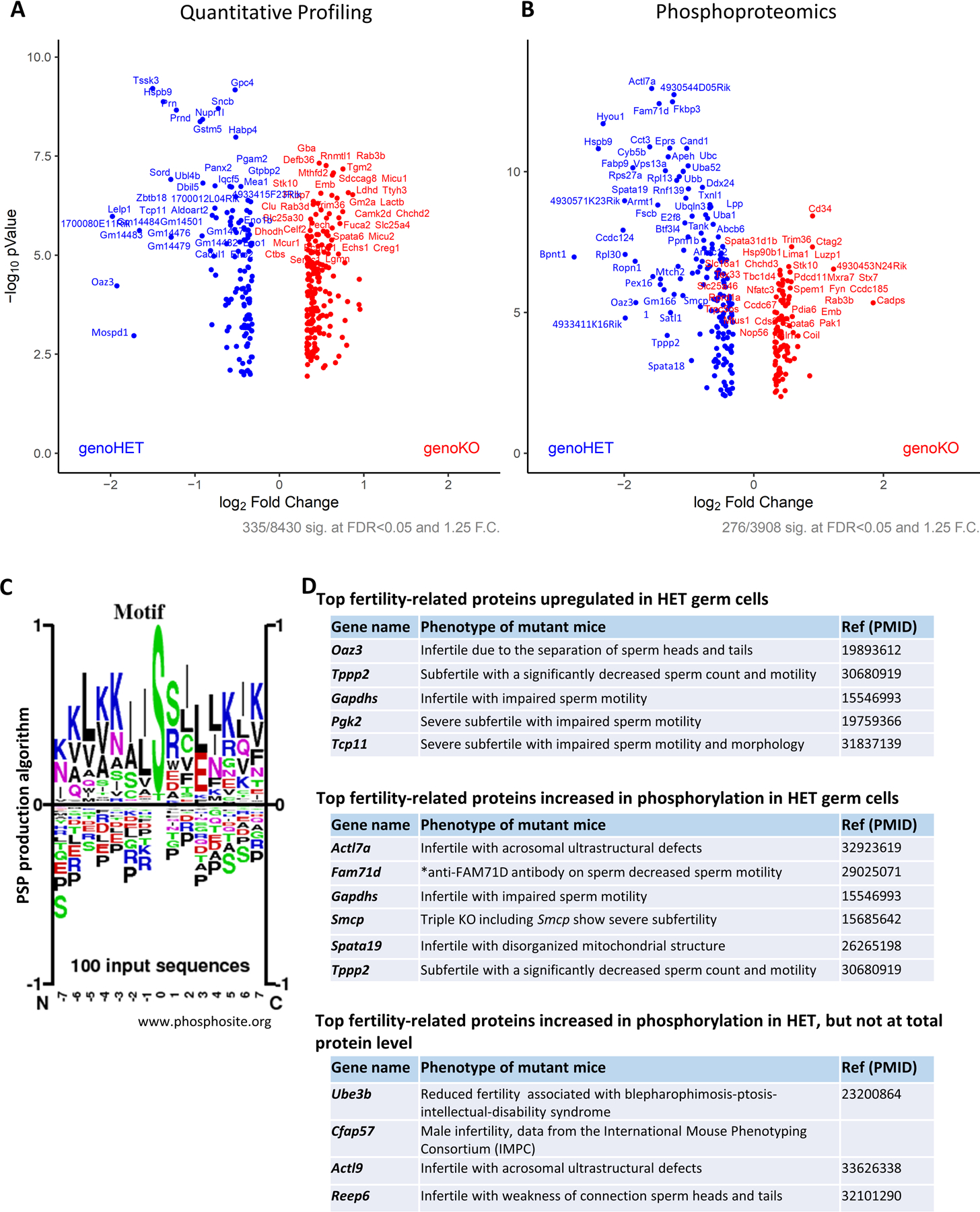
Results: TSSK3 is expressed in elongating sperm and localizes to the sperm tail. To define the essential roles of TSSK3 in vivo, heterozygous (HET) or homozygous KO male mice were mated with wild-type females, and fertility was assessed over 4 months; Tssk3 KO males are sterile, whereas HET males produced normal litter sizes. The absence of TSSK3 results in disorganization of all stages of testicular seminiferous epithelium and significantly increased vacuolization of germ cells, leading to dramatically reduced sperm counts and abnormal sperm morphology; despite these histologic changes, Tssk3 null mice have normal testis size. To elucidate the mechanisms causing the KO phenotype, we conducted phosphoproteomics using purified germ cells from Tssk3 HET and KO testes. We found that proteins implicated in male infertility, such as GAPDHS, ACTL7A, ACTL9, and REEP6, showed significantly reduced phosphorylation in KO testes compared to HET testes, despite unaltered total protein levels.
Conclusions: We demonstrated that TSSK3 is essential for male fertility and crucial for phosphorylation of multiple infertility-related proteins. These studies and the pathways in which TSSK3 functions have implications for human male infertility and nonhormonal contraception.
Keywords: knockout mouse; oligoteratozoospermia; oligozoospermia; proteomics; teratozoospermia.
© 2022 The Authors. Andrology published by Wiley Periodicals LLC on behalf of American Society of Andrology and European Academy of Andrology.
Figures
Similar articles
-
TSSK3, a novel target for male contraception, is required for spermiogenesis.Mol Reprod Dev. 2021 Nov;88(11):718-730. doi: 10.1002/mrd.23539. Epub 2021 Oct 8. Mol Reprod Dev. 2021. PMID: 34623009 Free PMC article.
-
Identification of TSSK1 and TSSK2 as Novel Targets for Male Contraception.Biomolecules. 2025 Apr 18;15(4):601. doi: 10.3390/biom15040601. Biomolecules. 2025. PMID: 40305308 Free PMC article.
-
Prss55 but not Prss51 is required for male fertility in mice†.Biol Reprod. 2020 Aug 4;103(2):223-234. doi: 10.1093/biolre/ioaa041. Biol Reprod. 2020. PMID: 32301961 Free PMC article.
-
Testis-specific serine kinase protein family in male fertility and as targets for non-hormonal male contraception†.Biol Reprod. 2020 Aug 4;103(2):264-274. doi: 10.1093/biolre/ioaa064. Biol Reprod. 2020. PMID: 32337545 Free PMC article. Review.
-
Androgen receptor roles in spermatogenesis and fertility: lessons from testicular cell-specific androgen receptor knockout mice.Endocr Rev. 2009 Apr;30(2):119-32. doi: 10.1210/er.2008-0025. Epub 2009 Jan 27. Endocr Rev. 2009. PMID: 19176467 Free PMC article. Review.
Cited by
-
Broad phosphorylation mediated by testis-specific serine/threonine kinases contributes to spermiogenesis and male fertility.Nat Commun. 2023 May 6;14(1):2629. doi: 10.1038/s41467-023-38357-0. Nat Commun. 2023. PMID: 37149634 Free PMC article.
-
Characterization of sexual maturity-associated N6-methyladenosine in boar testes.BMC Genomics. 2024 May 7;25(1):447. doi: 10.1186/s12864-024-10343-w. BMC Genomics. 2024. PMID: 38714941 Free PMC article.
-
Exposure to plastic debris alters expression of biomineralization, immune, and stress-related genes in the eastern oyster (Crassostrea virginica).PLoS One. 2025 Apr 29;20(4):e0319165. doi: 10.1371/journal.pone.0319165. eCollection 2025. PLoS One. 2025. PMID: 40299807 Free PMC article.
-
Structural specializations of the sperm tail.Cell. 2023 Jun 22;186(13):2880-2896.e17. doi: 10.1016/j.cell.2023.05.026. Epub 2023 Jun 15. Cell. 2023. PMID: 37327785 Free PMC article.
-
The role of HnrnpF/H as a driver of oligoteratozoospermia.iScience. 2024 Jun 6;27(7):110198. doi: 10.1016/j.isci.2024.110198. eCollection 2024 Jul 19. iScience. 2024. PMID: 39092172 Free PMC article.
References
-
- Oud MS, Smits RM, Smith HE, Mastrorosa FK, Holt GS, Houston BJ, de Vries PF, Alobaidi BKS, Batty LE, Ismail H, Greenwood J, Sheth H, Mikulasova A, Astuti GDN, Gilissen C, McEleny K, Turner H, Coxhead J, Cockell S, Braat DDM, Fleischer K, D’Hauwers KWM, Schaafsma E, Genetics of Male Infertility Initiative c, Nagirnaja L, Conrad DF, Friedrich C, Kliesch S, Aston KI, A Riera-Escamilla, C Krausz, Gonzaga-Jauregui C, Santibanez-Koref M, Elliott DJ, Vissers L, Tuttelmann F, O’Bryan MK, Ramos L, Xavier MJ, van der Heijden GW, Veltman JA. A de novo paradigm for male infertility. Nat Commun 2022; 13(1): 154. 10.1038/s41467-021-27132-8 - DOI - PMC - PubMed
-
- Houston BJ, Riera-Escamilla A, Wyrwoll MJ, Salas-Huetos A, Xavier MJ, Nagirnaja L, Friedrich C, Conrad DF, Aston KI, Krausz C, Tuttelmann F, O’Bryan MK, Veltman JA, Oud MS. A systematic review of the validated monogenic causes of human male infertility: 2020 update and a discussion of emerging gene-disease relationships. Hum Reprod Update 2021; 28(1): 15–29. 10.1093/humupd/dmab030 - DOI - PMC - PubMed
-
- Patel B, Parets S, Akana M, Kellogg G, Jansen M, Chang C, Cai Y, Fox R, Niknazar M, Shraga R, Hunter C, Pollock A, Wisotzkey R, Jaremko M, Bisignano A, Puig O. Comprehensive genetic testing for female and male infertility using next-generation sequencing. J Assist Reprod Genet 2018; 35(8): 1489–1496. 10.1007/s10815-018-1204-7 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
