

Neurexin1α knockout rats display oscillatory abnormalities and sensory processing deficits back-translating key endophenotypes of psychiatric disorders
- PMID: 36307390
- PMCID: PMC9616904
- DOI: 10.1038/s41398-022-02224-1
Neurexin1α knockout rats display oscillatory abnormalities and sensory processing deficits back-translating key endophenotypes of psychiatric disorders
Abstract
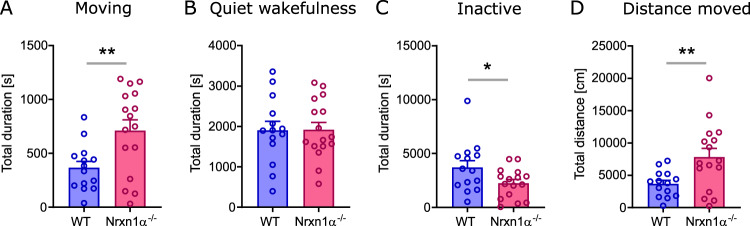
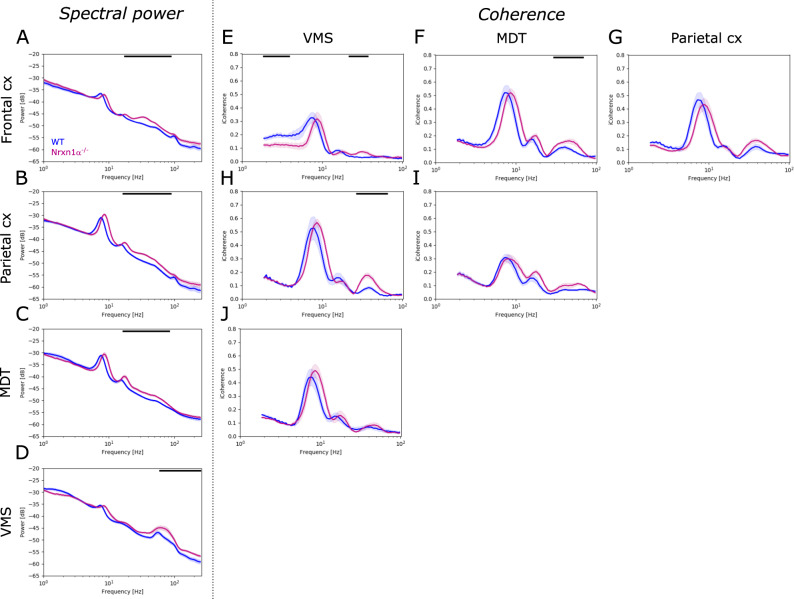
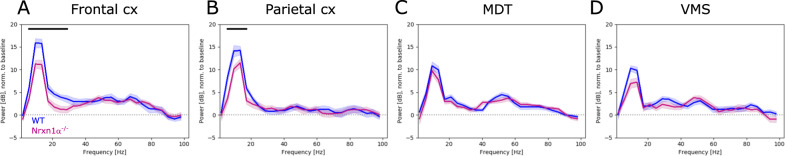
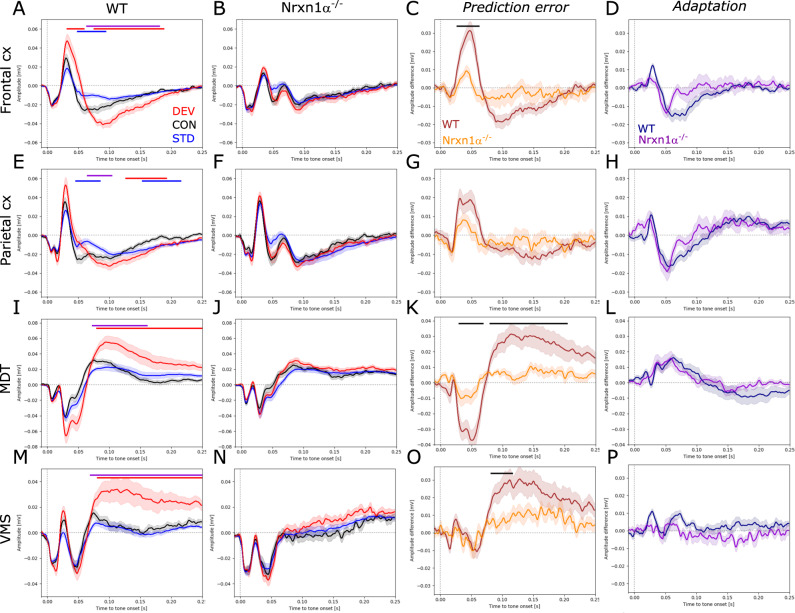
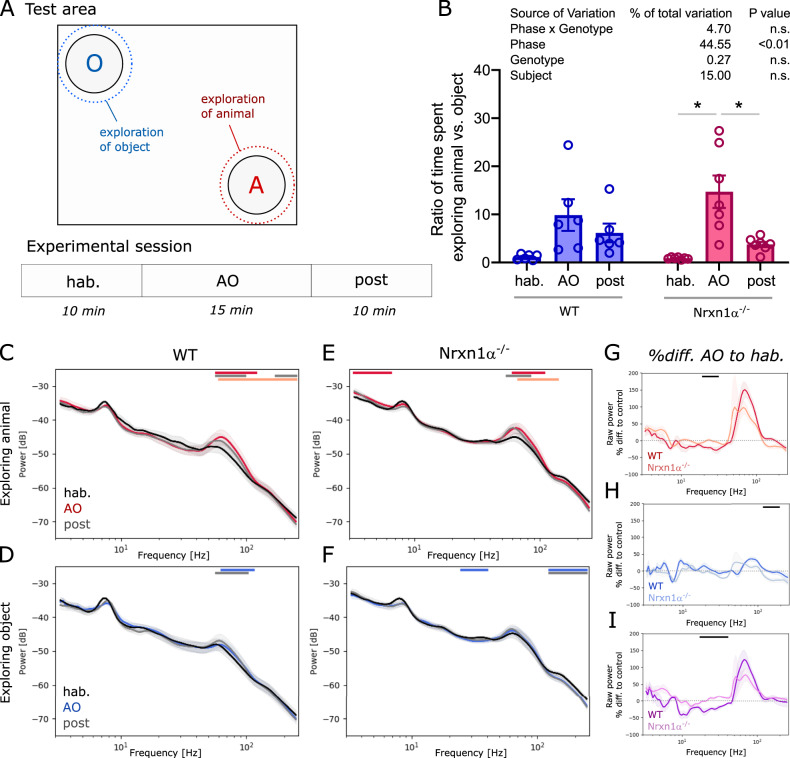
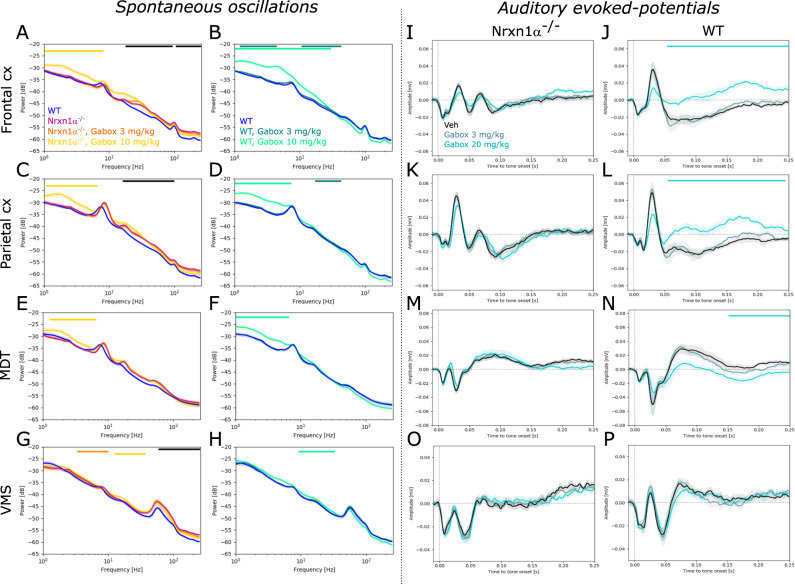
Neurexins are presynaptic transmembrane proteins crucial for synapse development and organization. Deletion and missense mutations in all three Neurexin genes have been identified in psychiatric disorders, with mutations in the NRXN1 gene most strongly linked to schizophrenia (SZ) and autism spectrum disorder (ASD). While the consequences of NRXN1 deletion have been extensively studied on the synaptic and behavioral levels, circuit endophenotypes that translate to the human condition have not been characterized yet. Therefore, we investigated the electrophysiology of cortico-striatal-thalamic circuits in Nrxn1α-/- rats and wildtype littermates focusing on a set of translational readouts, including spontaneous oscillatory activity, auditory-evoked oscillations and potentials, as well as mismatch negativity-like (MMN) responses and responses to social stimuli. On the behavioral level Nrxn1α-/- rats showed locomotor hyperactivity. In vivo freely moving electrophysiology revealed pronounced increases of spontaneous oscillatory power within the gamma band in all studied brain areas and elevation of gamma coherence in cortico-striatal and thalamocortical circuits of Nrxn1α-/- rats. In contrast, auditory-evoked oscillations driven by chirp-modulated tones showed reduced power in cortical areas confined to slower oscillations. Finally, Nrxn1α-/- rats exhibited altered auditory evoked-potentials and profound deficits in MMN-like responses, explained by reduced prediction error. Despite deficits for auditory stimuli, responses to social stimuli appeared intact. A central hypothesis for psychiatric and neurodevelopmental disorders is that a disbalance of excitation-to-inhibition is underlying oscillatory and sensory deficits. In a first attempt to explore the impact of inhibitory circuit modulation, we assessed the effects of enhancing tonic inhibition via δ-containing GABAA receptors (using Gaboxadol) on endophenotypes possibly associated with network hyperexcitability. Pharmacological experiments applying Gaboxadol showed genotype-specific differences, but failed to normalize oscillatory or sensory processing abnormalities. In conclusion, our study revealed endophenotypes in Nrxn1α-/- rats that could be used as translational biomarkers for drug development in psychiatric disorders.
© 2022. The Author(s).
Conflict of interest statement
PJ, MB, SM, PS, and RLR were under paid employment by the company F. Hoffmann-La Roche (Roche). The funder provided support in the form of salaries for authors, but did not have any additional role in the study design, data collection and analysis, decision to publish, or preparation of the manuscript. This does not alter the authors’ adherence to all the journal policies on sharing data and materials.
Figures
References
-
- Missler M, Zhang W, Rohlmann A, Kattenstroth G, Hammer RE, Gottmann K, et al. Alpha-neurexins couple Ca2+ channels to synaptic vesicle exocytosis. Nature. 2003;423:939–48. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
