

Arabidopsis ABCG14 forms a homodimeric transporter for multiple cytokinins and mediates long-distance transport of isopentenyladenine-type cytokinins
- PMID: 36307987
- PMCID: PMC10030318
- DOI: 10.1016/j.xplc.2022.100468
Arabidopsis ABCG14 forms a homodimeric transporter for multiple cytokinins and mediates long-distance transport of isopentenyladenine-type cytokinins
Abstract
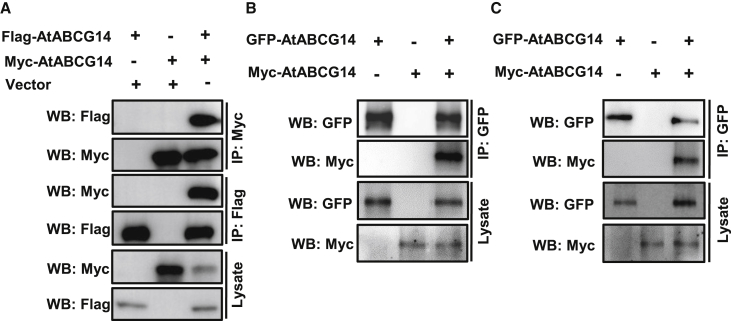
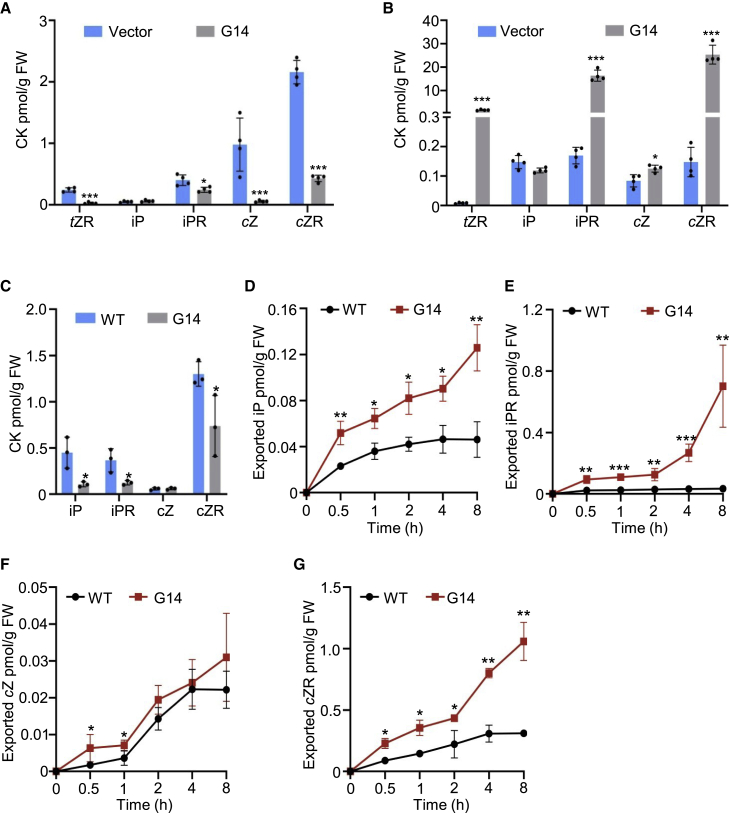
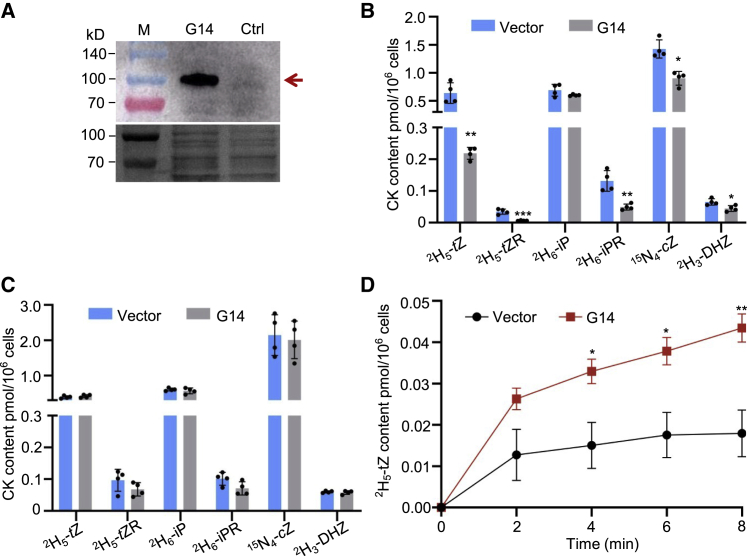
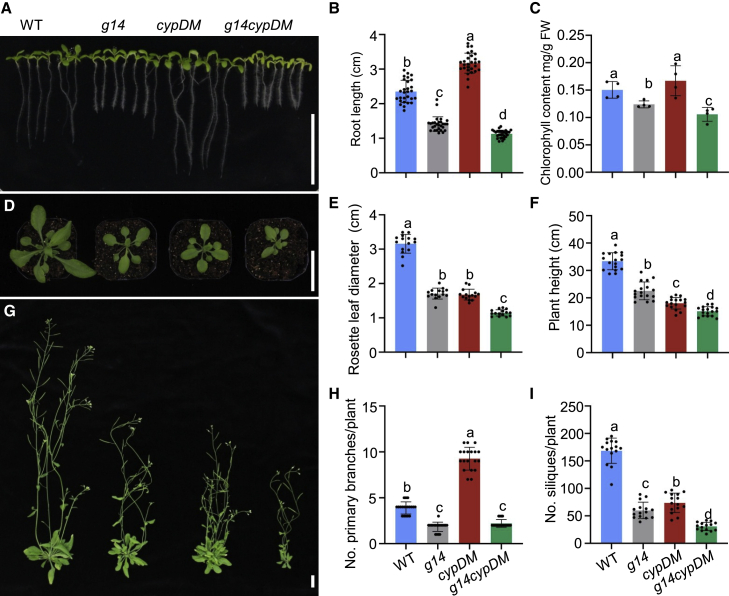
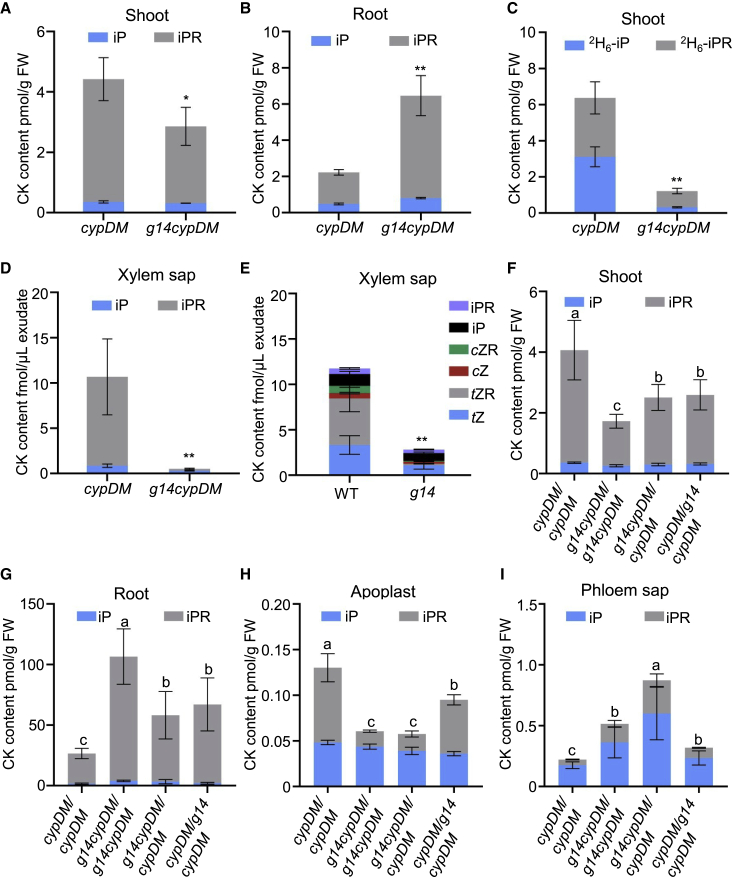
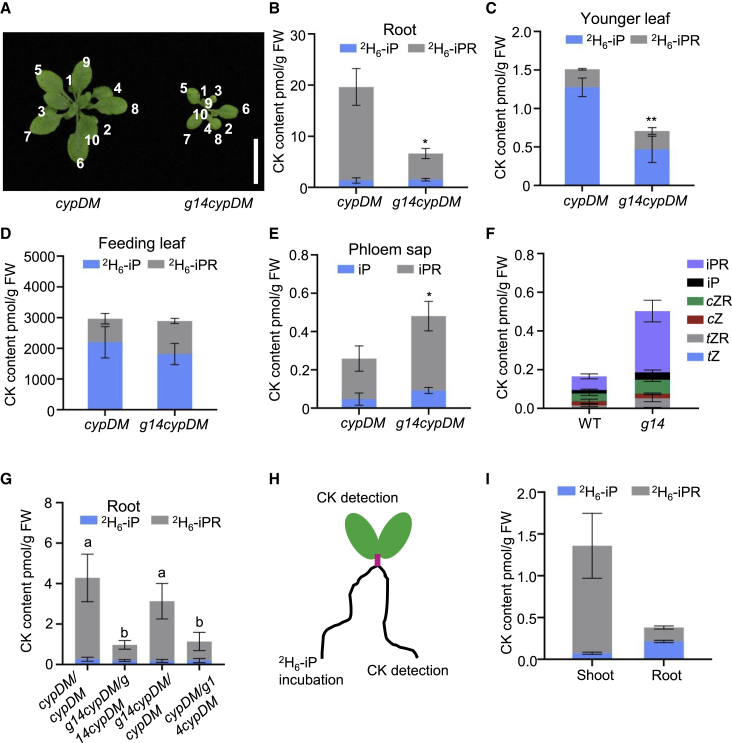
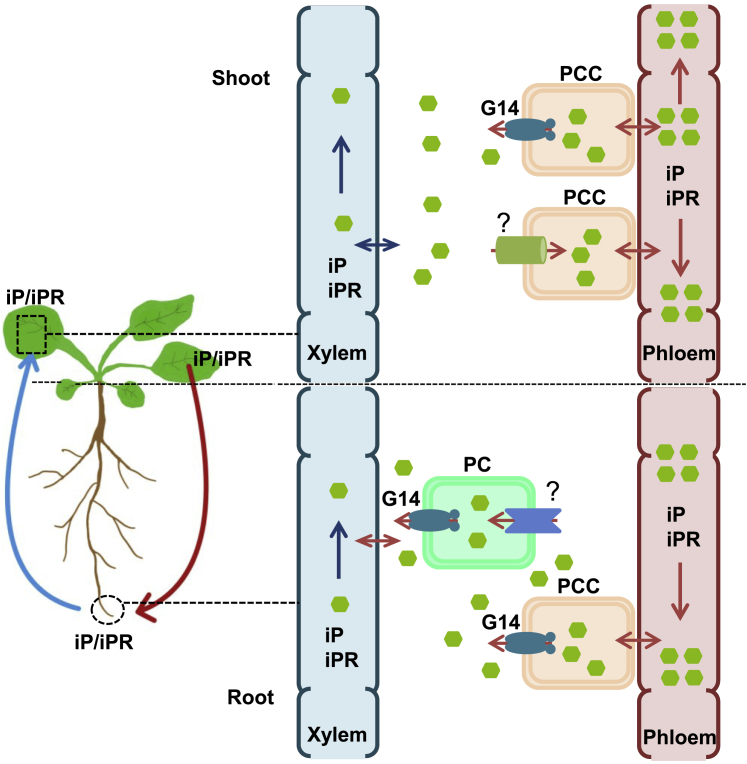
Cytokinins (CKs), primarily trans-zeatin (tZ) and isopentenyladenine (iP) types, play critical roles in plant growth, development, and various stress responses. Long-distance transport of tZ-type CKs meidated by Arabidopsis ATP-binding cassette transporter subfamily G14 (AtABCG14) has been well studied; however, less is known about the biochemical properties of AtABCG14 and its transporter activity toward iP-type CKs. Here we reveal the biochemical properties of AtABCG14 and provide evidence that it is also required for long-distance transport of iP-type CKs. AtABCG14 formed homodimers in human (Homo sapiens) HEK293T, tobacco (Nicotiana tabacum), and Arabidopsis cells. Transporter activity assays of AtABCG14 in Arabidopsis, tobacco, and yeast (Saccharomyces cerevisiae) showed that AtABCG14 may directly transport multiple CKs, including iP- and tZ-type species. AtABCG14 expression was induced by iP in a tZ-type CK-deficient double mutant (cypDM) of CYP735A1 and CYP735A2. The atabcg14 cypDM triple mutant exhibited stronger CK-deficiency phenotypes than cypDM. Hormone profiling, reciprocal grafting, and 2H6-iP isotope tracer experiments showed that root-to-shoot and shoot-to-root long-distance transport of iP-type CKs were suppressed in atabcg14 cypDM and atabcg14. These results suggest that AtABCG14 participates in three steps of the circular long-distance transport of iP-type CKs: xylem loading in the root for shootward transport, phloem unloading in the shoot for shoot distribution, and phloem unloading in the root for root distribution. We found that AtABCG14 displays transporter activity toward multiple CK species and revealed its versatile roles in circular long-distance transport of iP-type CKs. These findings provide new insights into the transport mechanisms of CKs and other plant hormones.
Keywords: circular transport; half-size ABC transporter; homodimer; isopentenyladenine-type cytokinins; long-distance transport.
Copyright © 2022 The Author(s). Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Ashikari M., Sakakibara H., Lin S., Yamamoto T., Takashi T., Nishimura A., Angeles E.R., Qian Q., Kitano H., Matsuoka M. Cytokinin oxidase regulates rice grain production. Science. 2005;309:741–745. - PubMed
-
- Banasiak J., Jasiński M. ATP-binding cassette transporters in nonmodel plants. New Phytol. 2021;233:1597–1612. - PubMed
-
- Bishopp A., Lehesranta S., Vatén A., Help H., El-Showk S., Scheres B., Helariutta K., Mähönen A.P., Sakakibara H., Helariutta Y. Phloem-transported cytokinin regulates polar auxin transport and maintains vascular pattern in the root meristem. Curr. Biol. 2011;21:927–932. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
