

Rieske Oxygenase Catalyzed C-H Bond Functionalization Reactions in Chlorophyll b Biosynthesis
- PMID: 36313167
- PMCID: PMC9615114
- DOI: 10.1021/acscentsci.2c00058
Rieske Oxygenase Catalyzed C-H Bond Functionalization Reactions in Chlorophyll b Biosynthesis
Abstract
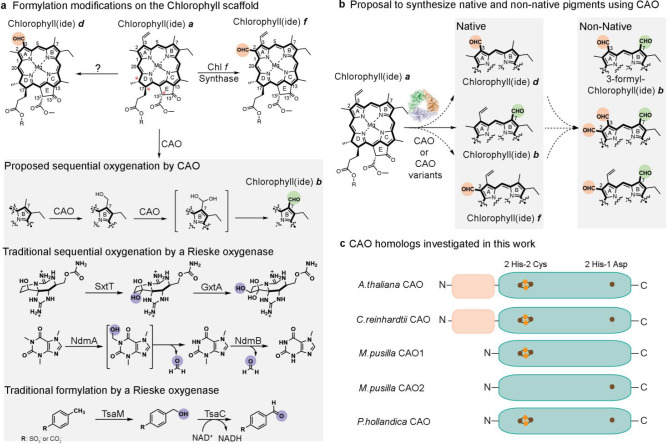
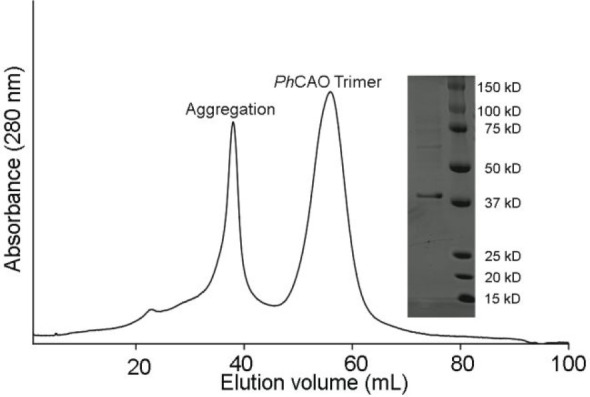
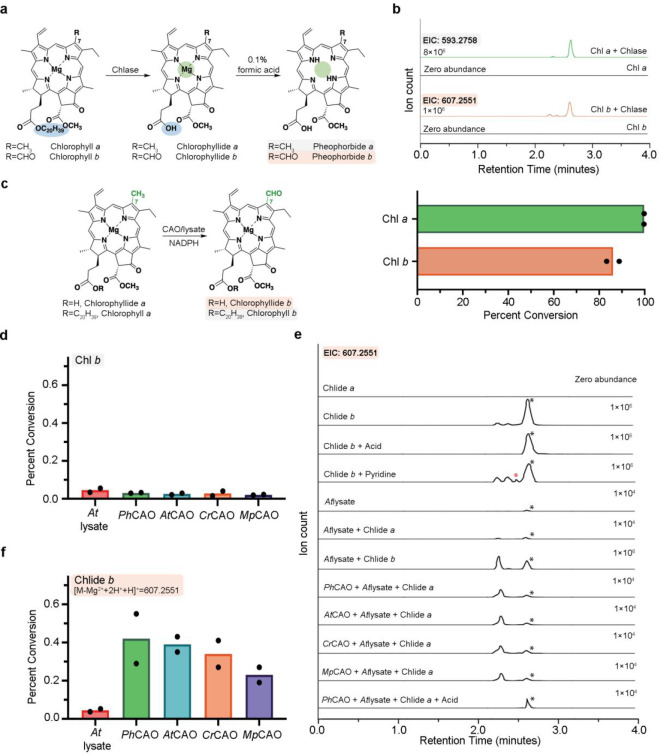
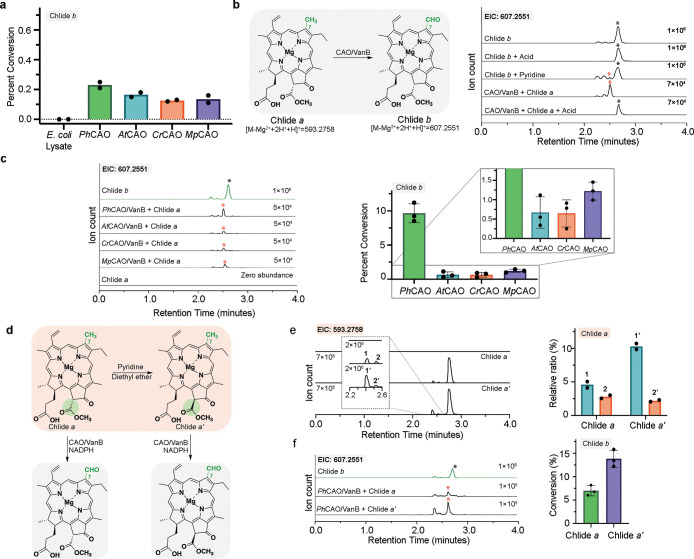
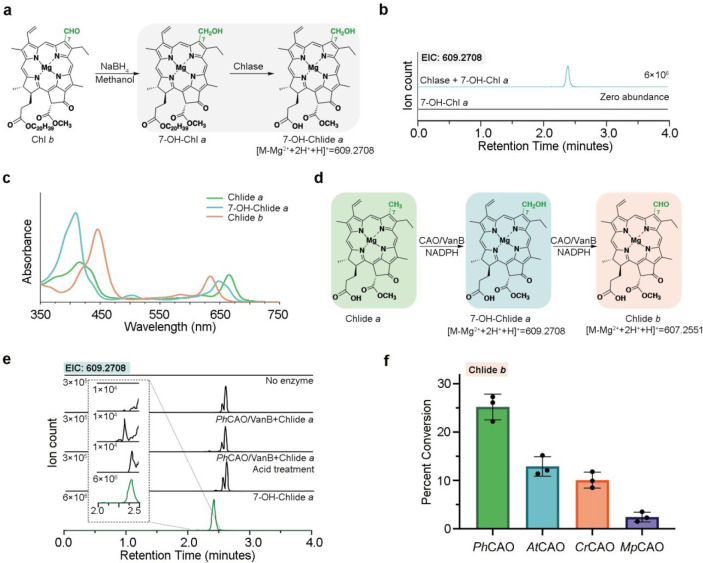
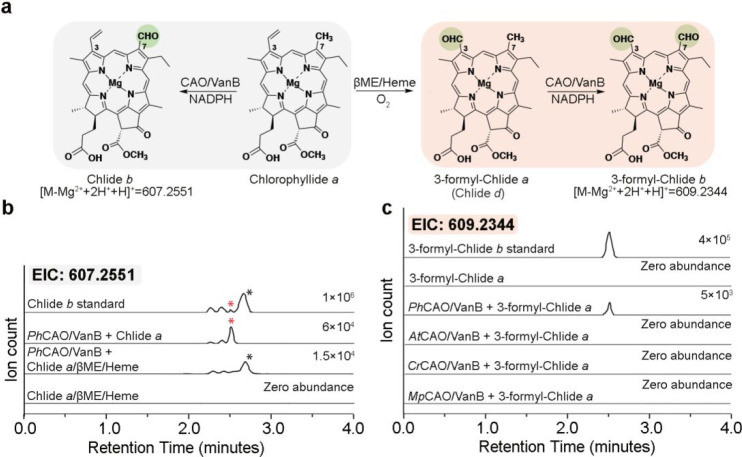
Rieske oxygenases perform precise C-H bond functionalization reactions in anabolic and catabolic pathways. These reactions are typically characterized as monooxygenation or dioxygenation reactions, but other divergent reactions are also catalyzed by Rieske oxygenases. Chlorophyll(ide) a oxygenase (CAO), for example is proposed to catalyze two monooxygenation reactions to transform a methyl-group into the formyl-group of Chlorophyll b. This formyl group, like the formyl groups found in other chlorophyll pigments, tunes the absorption spectra of chlorophyllb and supports the ability of several photosynthetic organisms to adapt to environmental light. Despite the importance of this reaction, CAO has never been studied in vitro with purified protein, leaving many open questions regarding whether CAO can facilitate both oxygenation reactions using just the Rieske oxygenase machinery. In this study, we demonstrated that four CAO homologues in partnership with a non-native reductase convert a Chlorophyll a precursor, chlorophyllidea, into chlorophyllideb in vitro. Analysis of this reaction confirmed the existence of the proposed intermediate, highlighted the stereospecificity of the reaction, and revealed the potential of CAO as a tool for synthesizing custom-tuned natural and unnatural chlorophyll pigments. This work thus adds to our fundamental understanding of chlorophyll biosynthesis and Rieske oxygenase chemistry.
© 2022 The Authors. Published by American Chemical Society.
Conflict of interest statement
The authors declare no competing financial interest.
Figures
References
LinkOut - more resources
Full Text Sources