

WTAP Targets the METTL3 m6A-Methyltransferase Complex to Cytoplasmic Hepatitis C Virus RNA to Regulate Infection
- PMID: 36314819
- PMCID: PMC9683008
- DOI: 10.1128/jvi.00997-22
WTAP Targets the METTL3 m6A-Methyltransferase Complex to Cytoplasmic Hepatitis C Virus RNA to Regulate Infection
Abstract
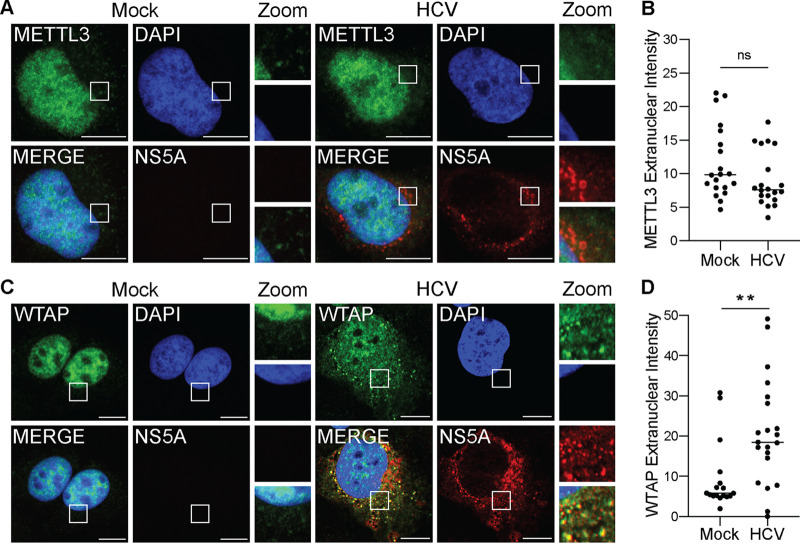
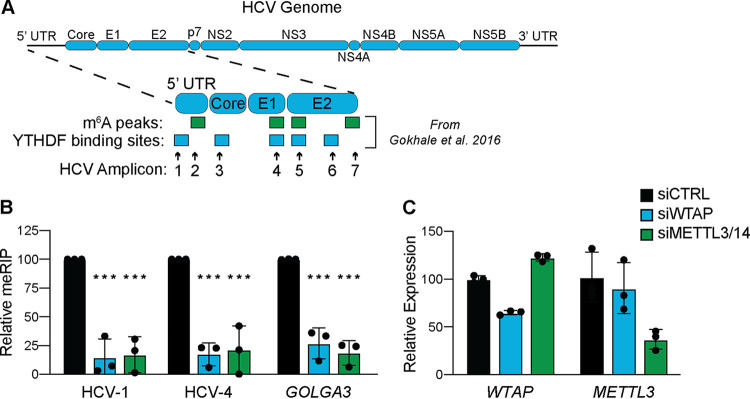
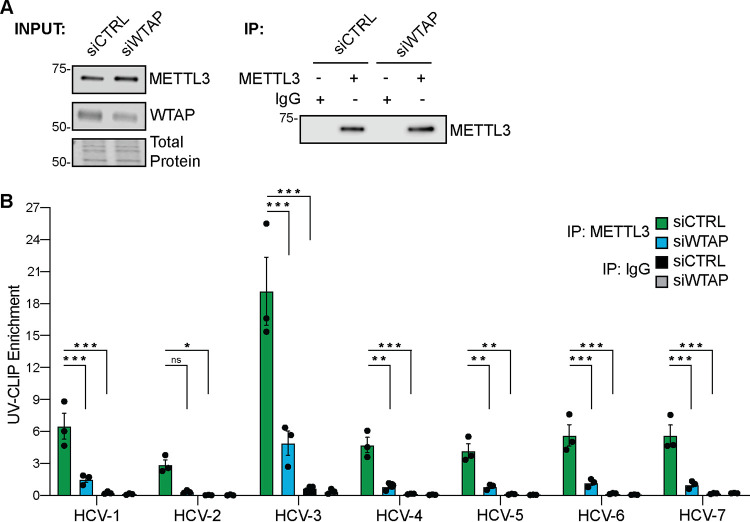
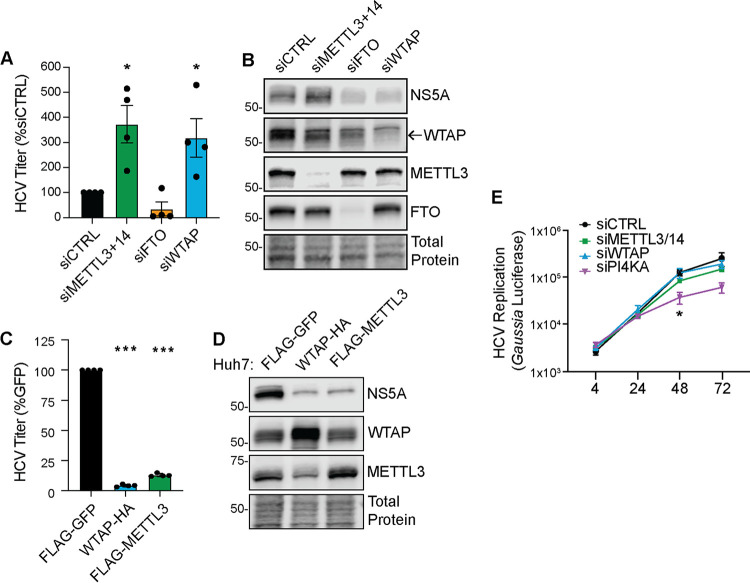
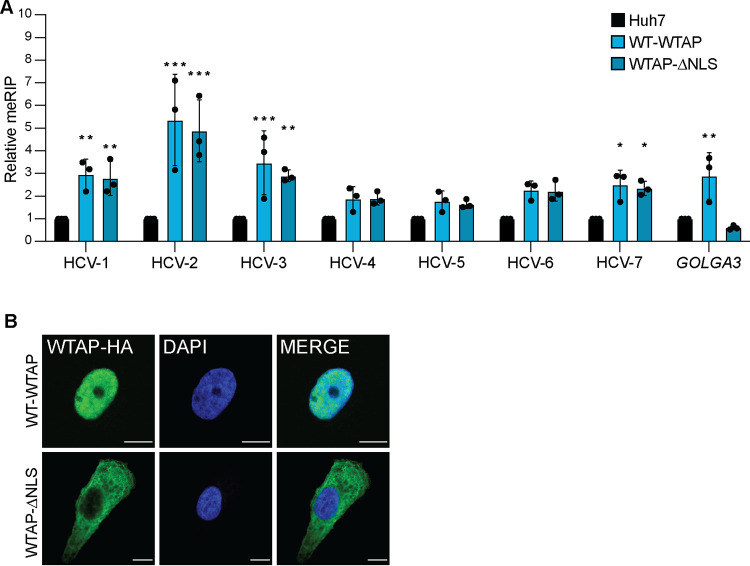
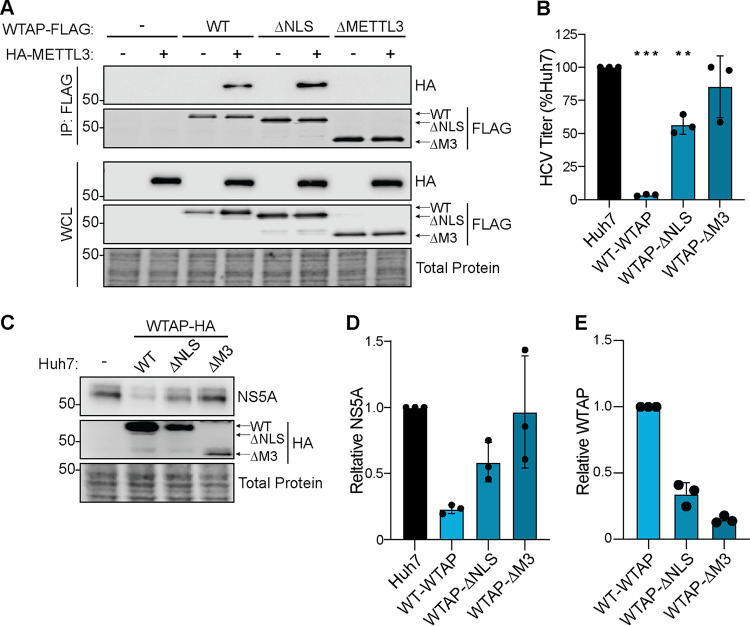
Modification of the hepatitis C virus (HCV) positive-strand RNA genome by N6-methyladenosine (m6A) regulates the viral life cycle. This life cycle takes place solely in the cytoplasm, while m6A addition on cellular mRNA takes place in the nucleus. Thus, the mechanisms by which m6A is deposited on the viral RNA have been unclear. In this work, we find that m6A modification of HCV RNA by the m6A-methyltransferase proteins methyltransferase-like 3 and 14 (METTL3 and METTL14) is regulated by Wilms' tumor 1-associating protein (WTAP). WTAP, a predominantly nuclear protein, is an essential member of the cellular mRNA m6A-methyltransferase complex and known to target METTL3 to mRNA. We found that HCV infection induces localization of WTAP to the cytoplasm. Importantly, we found that WTAP is required for both METTL3 interaction with HCV RNA and m6A modification across the viral RNA genome. Further, we found that WTAP, like METTL3 and METTL14, negatively regulates the production of infectious HCV virions, a process that we have previously shown is regulated by m6A. Excitingly, WTAP regulation of both HCV RNA m6A modification and virion production was independent of its ability to localize to the nucleus. Together, these results reveal that WTAP is critical for HCV RNA m6A modification by METTL3 and METTL14 in the cytoplasm. IMPORTANCE Positive-strand RNA viruses such as HCV represent a significant global health burden. Previous work has described that HCV RNA contains the RNA modification m6A and how this modification regulates viral infection. Yet, how this modification is targeted to HCV RNA has remained unclear due to the incompatibility of the nuclear cellular processes that drive m6A modification with the cytoplasmic HCV life cycle. In this study, we present evidence for how m6A modification is targeted to HCV RNA in the cytoplasm by a mechanism in which WTAP recruits the m6A-methyltransferase METTL3 to HCV RNA. This targeting strategy for m6A modification of cytoplasmic RNA viruses is likely relevant for other m6A-modified positive-strand RNA viruses with cytoplasmic life cycles such as enterovirus 71 and SARS-CoV-2 and provides an exciting new target for potential antiviral therapies.
Keywords: HCV; METTL14; METTL3; N6-methyladenosine; RNA modification; WTAP.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Update of
-
WTAP targets the METTL3 m 6 A-methyltransferase complex to cytoplasmic hepatitis C virus RNA to regulate infection.bioRxiv [Preprint]. 2022 Jun 28:2022.06.27.497872. doi: 10.1101/2022.06.27.497872. bioRxiv. 2022. Update in: J Virol. 2022 Nov 23;96(22):e0099722. doi: 10.1128/jvi.00997-22. PMID: 35794896 Free PMC article. Updated. Preprint.
References
-
- Gokhale NS, McIntyre ABR, McFadden MJ, Roder AE, Kennedy EM, Gandara JA, Hopcraft SE, Quicke KM, Vazquez C, Willer J, Ilkayeva OR, Law BA, Holley CL, Garcia-Blanco MA, Evans MJ, Suthar MS, Bradrick SS, Mason CE, Horner SM. 2016. N6-methyladenosine in flaviviridae viral RNA genomes regulates infection. Cell Host Microbe 20:654–665. 10.1016/j.chom.2016.09.015. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
