

Bacteriophage genome engineering with CRISPR-Cas13a
- PMID: 36316452
- PMCID: PMC9722621
- DOI: 10.1038/s41564-022-01243-4
Bacteriophage genome engineering with CRISPR-Cas13a
Abstract
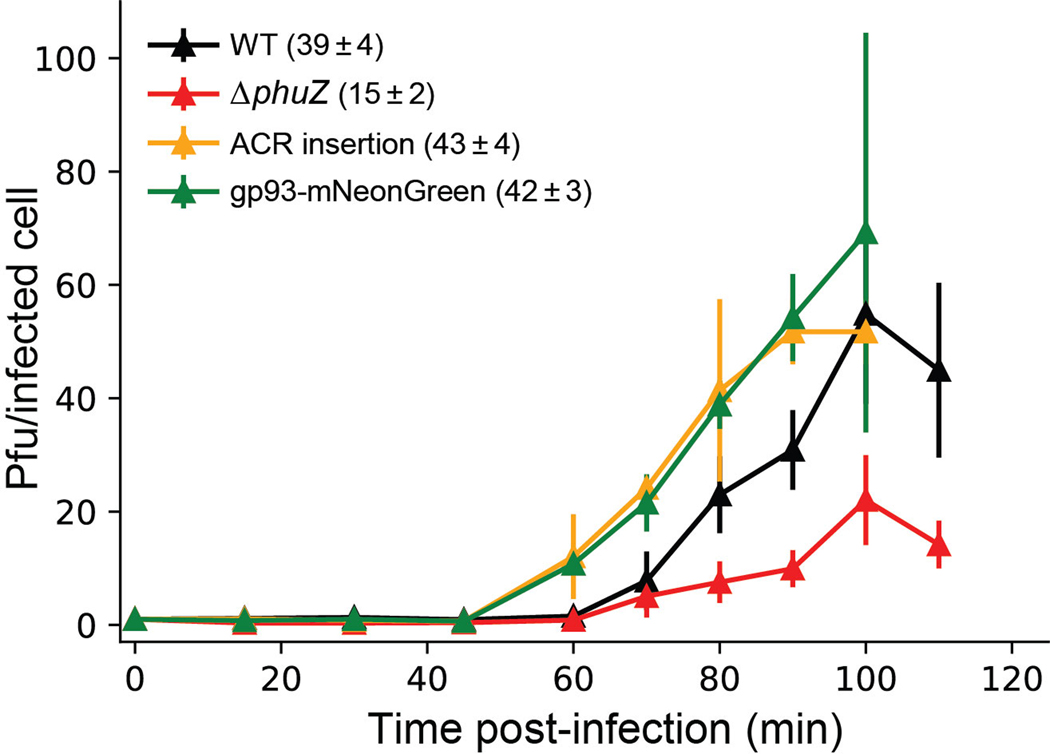
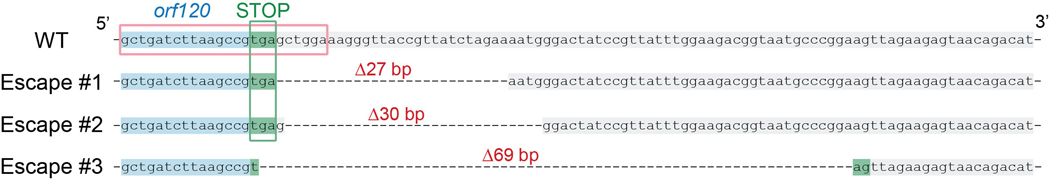
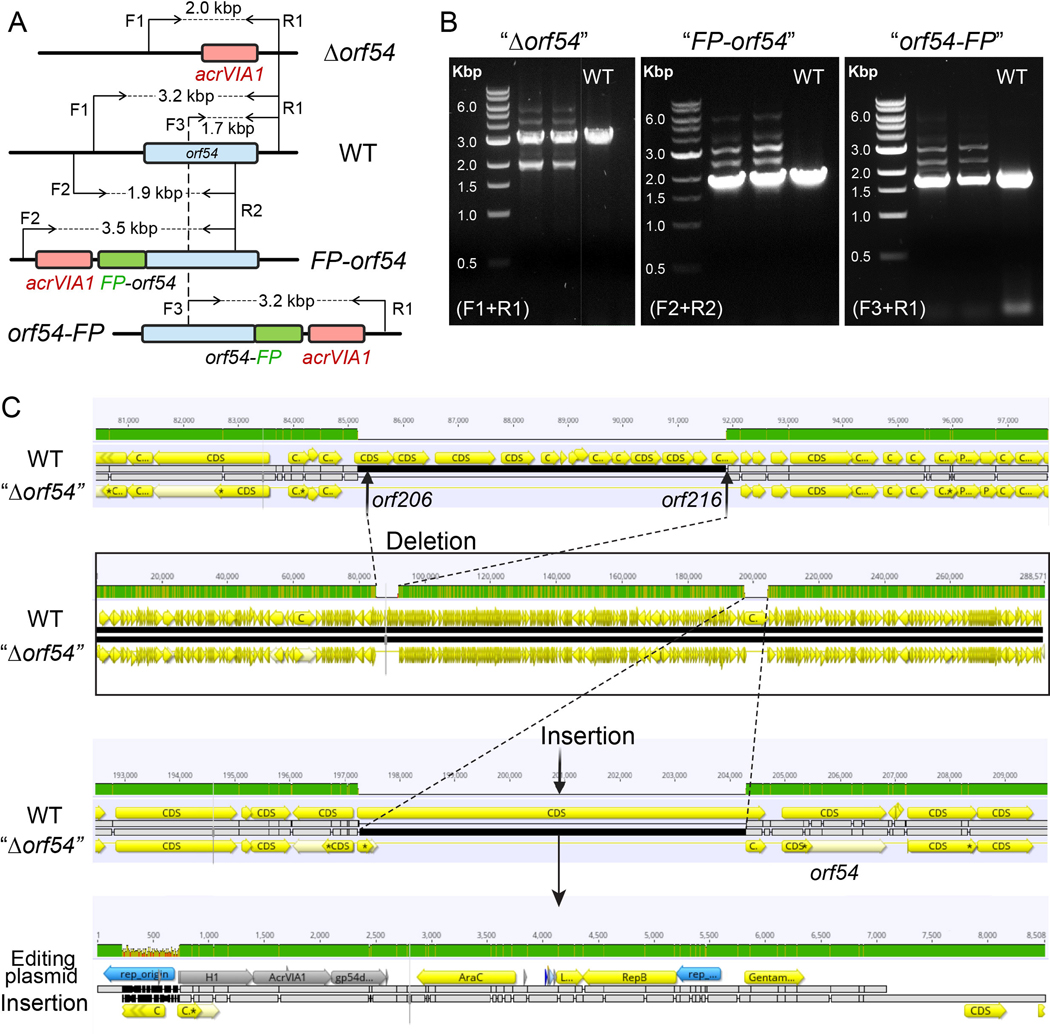
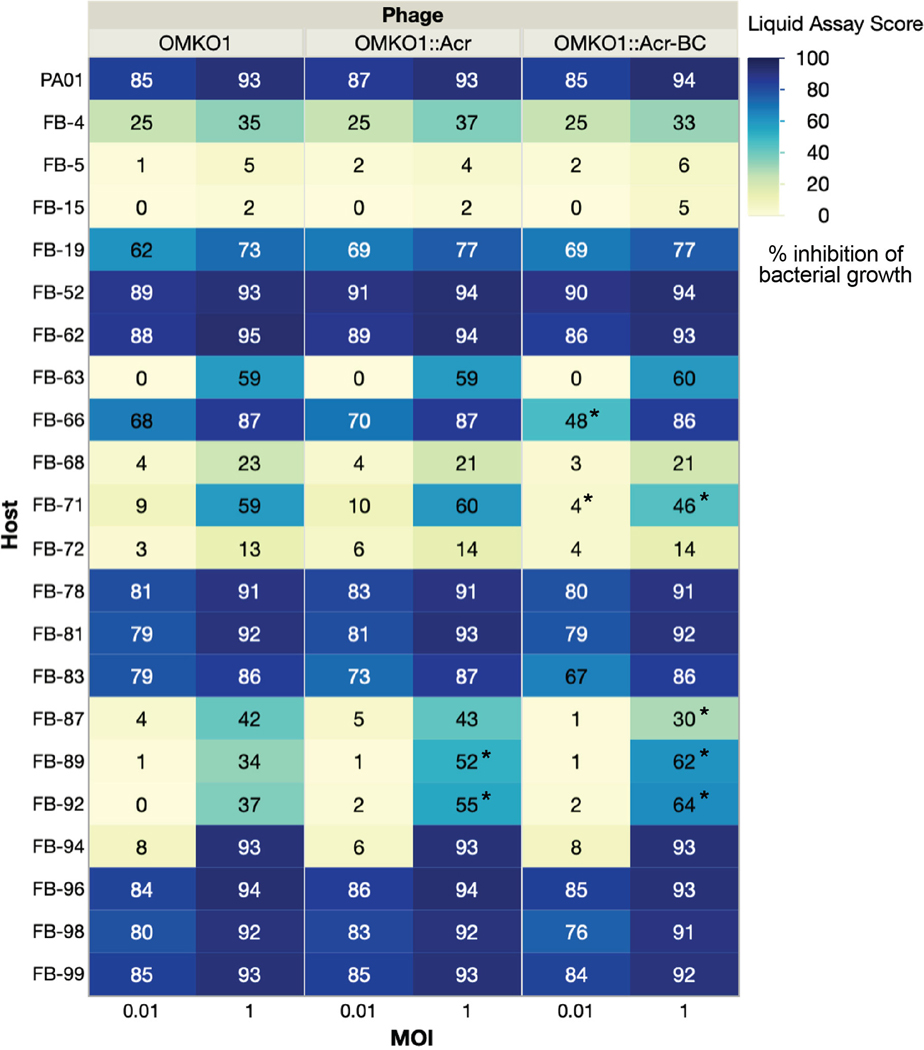
Jumbo phages such as Pseudomonas aeruginosa ФKZ have potential as antimicrobials and as a model for uncovering basic phage biology. Both pursuits are currently limited by a lack of genetic engineering tools due to a proteinaceous 'phage nucleus' structure that protects from DNA-targeting CRISPR-Cas tools. To provide reverse-genetics tools for DNA jumbo phages from this family, we combined homologous recombination with an RNA-targeting CRISPR-Cas13a enzyme and used an anti-CRISPR gene (acrVIA1) as a selectable marker. We showed that this process can insert foreign genes, delete genes and add fluorescent tags to genes in the ФKZ genome. Fluorescent tagging of endogenous gp93 revealed that it is ejected with the phage DNA while deletion of the tubulin-like protein PhuZ surprisingly had only a modest impact on phage burst size. Editing of two other phages that resist DNA-targeting CRISPR-Cas systems was also achieved. RNA-targeting Cas13a holds great promise for becoming a universal genetic editing tool for intractable phages, enabling the systematic study of phage genes of unknown function.
© 2022. The Author(s), under exclusive licence to Springer Nature Limited.
Figures


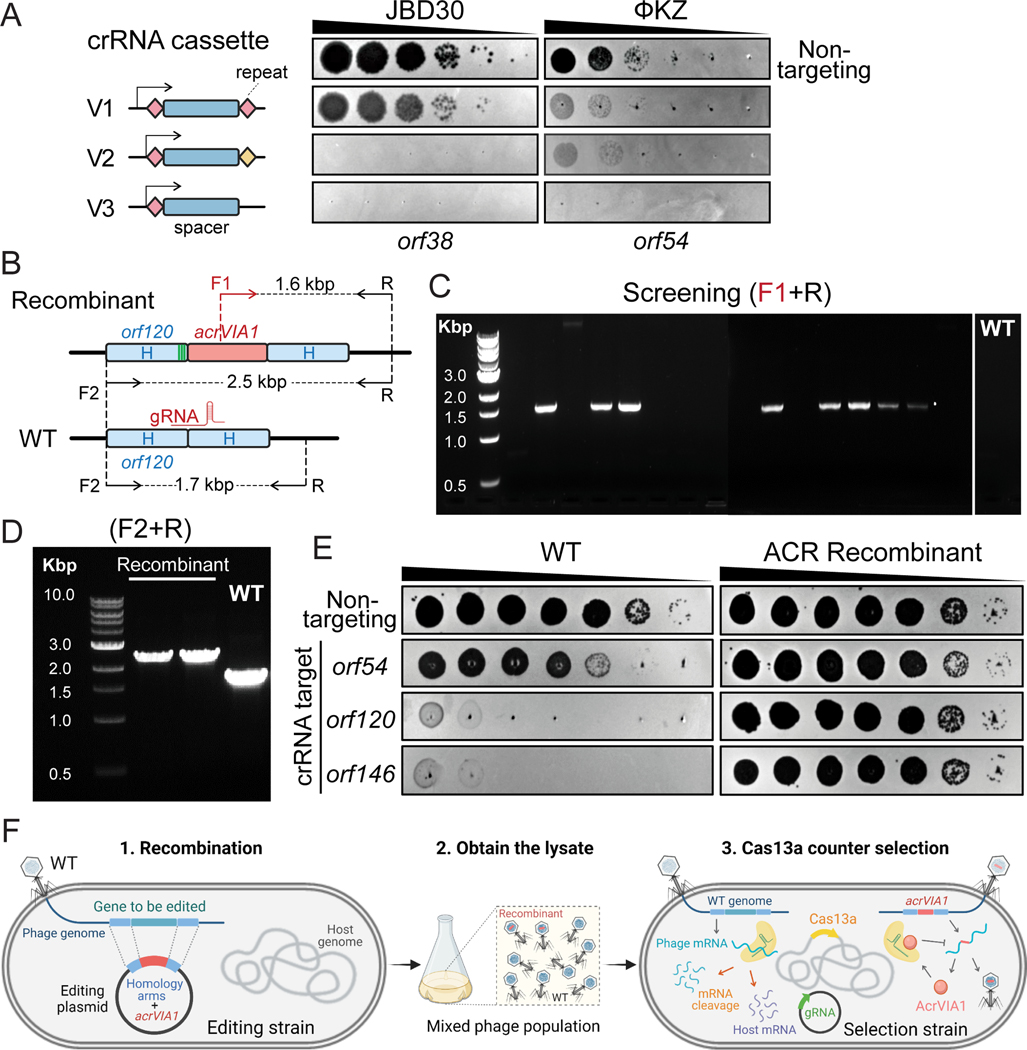
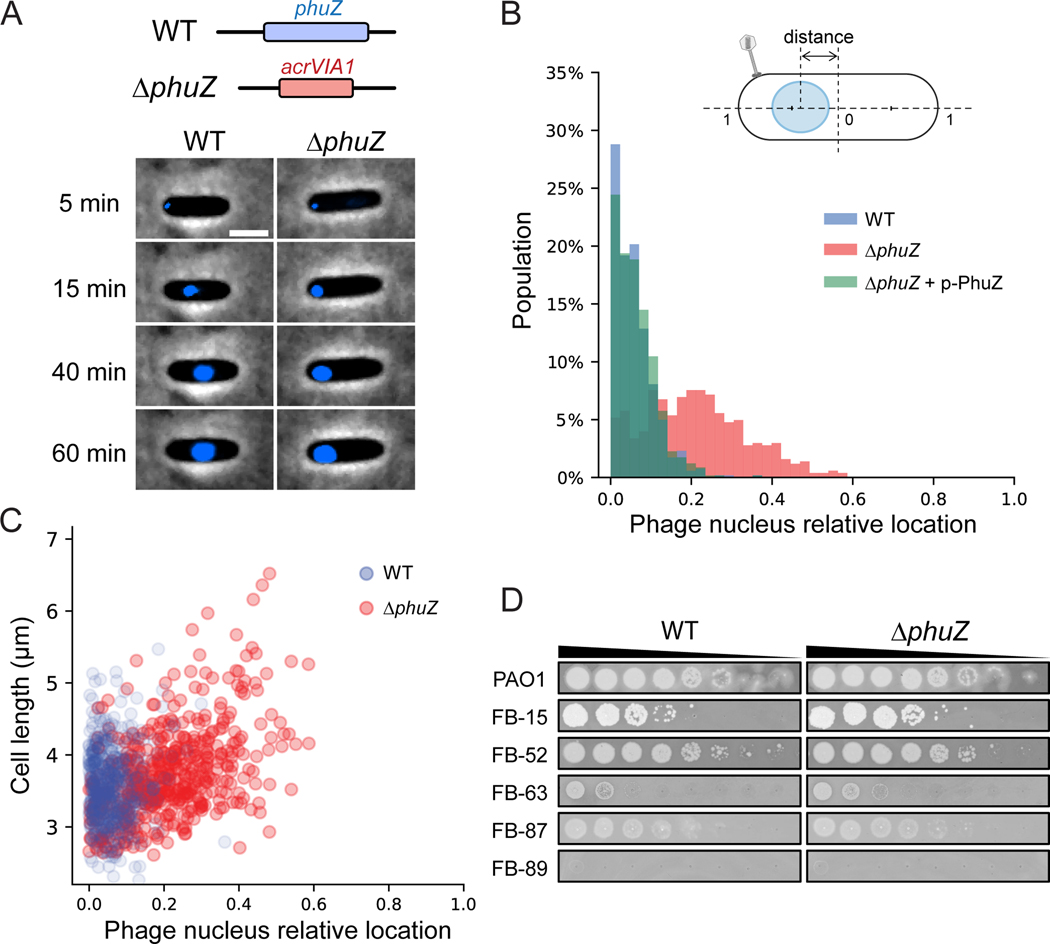
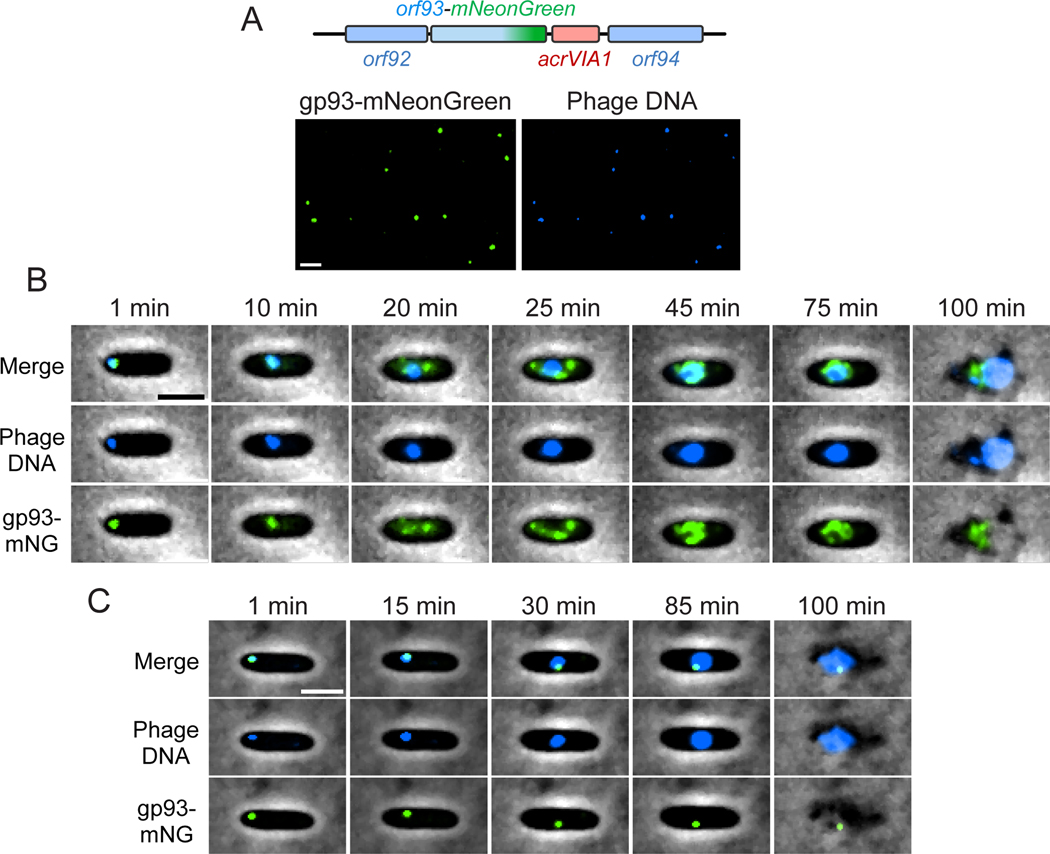
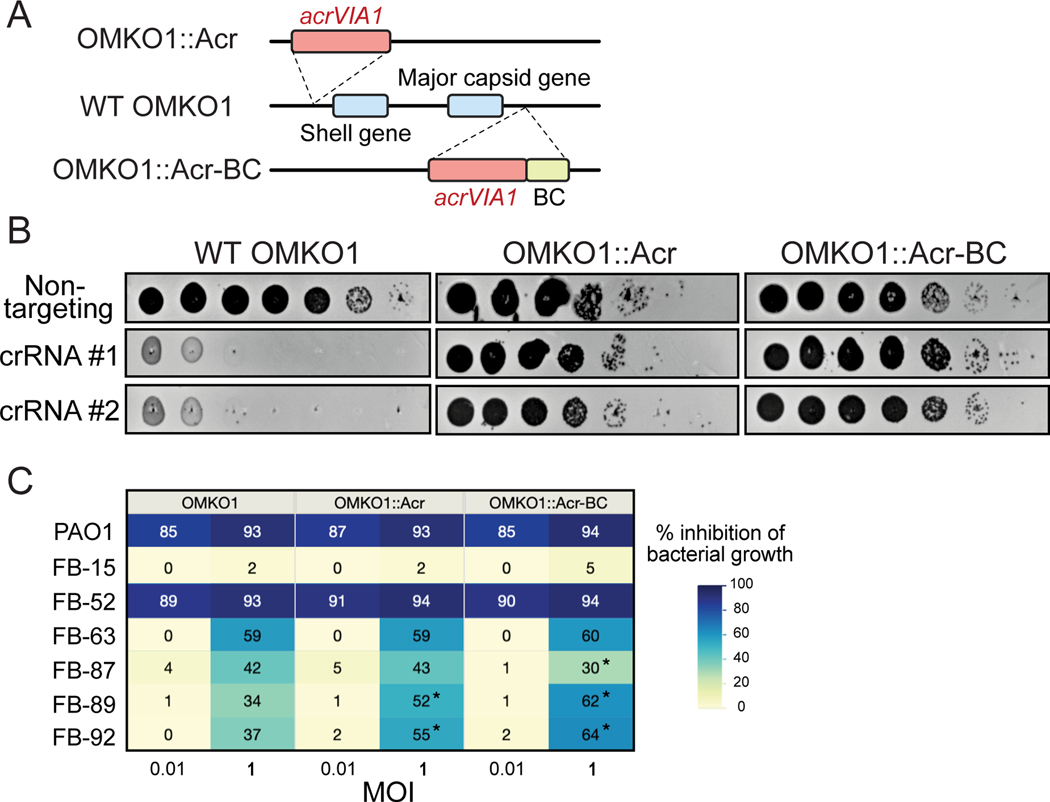
References
-
- Kortright KE, Chan BK, Koff JL, and Turner PE (2019). Phage Therapy: A Renewed Approach to Combat Antibiotic-Resistant Bacteria. Cell Host Microbe 25, 219–232. - PubMed
-
- Nobrega FL, Costa AR, Kluskens LD, and Azeredo J (2015). Revisiting phage therapy: new applications for old resources. Trends Microbiol 23, 185–191. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
