

Mitochondrial Uncoupling Induces Epigenome Remodeling and Promotes Differentiation in Neuroblastoma
- PMID: 36318118
- PMCID: PMC9851961
- DOI: 10.1158/0008-5472.CAN-22-1029
Mitochondrial Uncoupling Induces Epigenome Remodeling and Promotes Differentiation in Neuroblastoma
Erratum in
-
Correction: Mitochondrial Uncoupling Induces Epigenome Remodeling and Promotes Differentiation in Neuroblastoma.Cancer Res. 2023 Jan 18;83(2):346. doi: 10.1158/0008-5472.CAN-22-3873. Cancer Res. 2023. PMID: 36651077 No abstract available.
Abstract
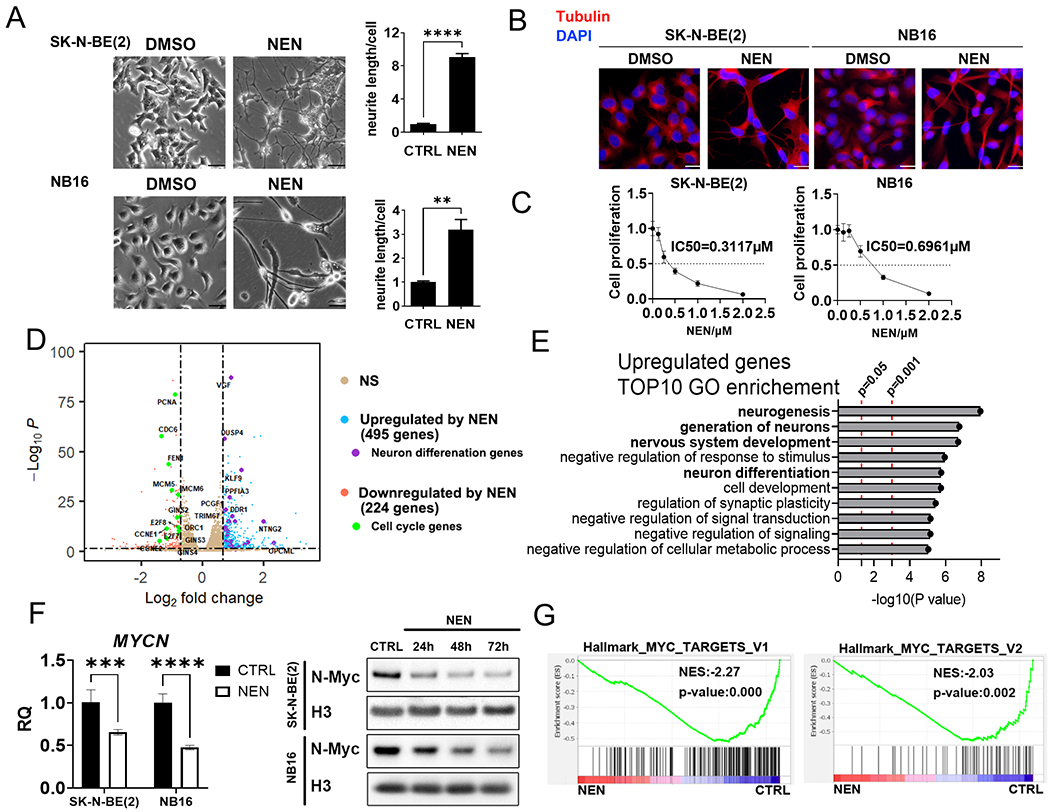
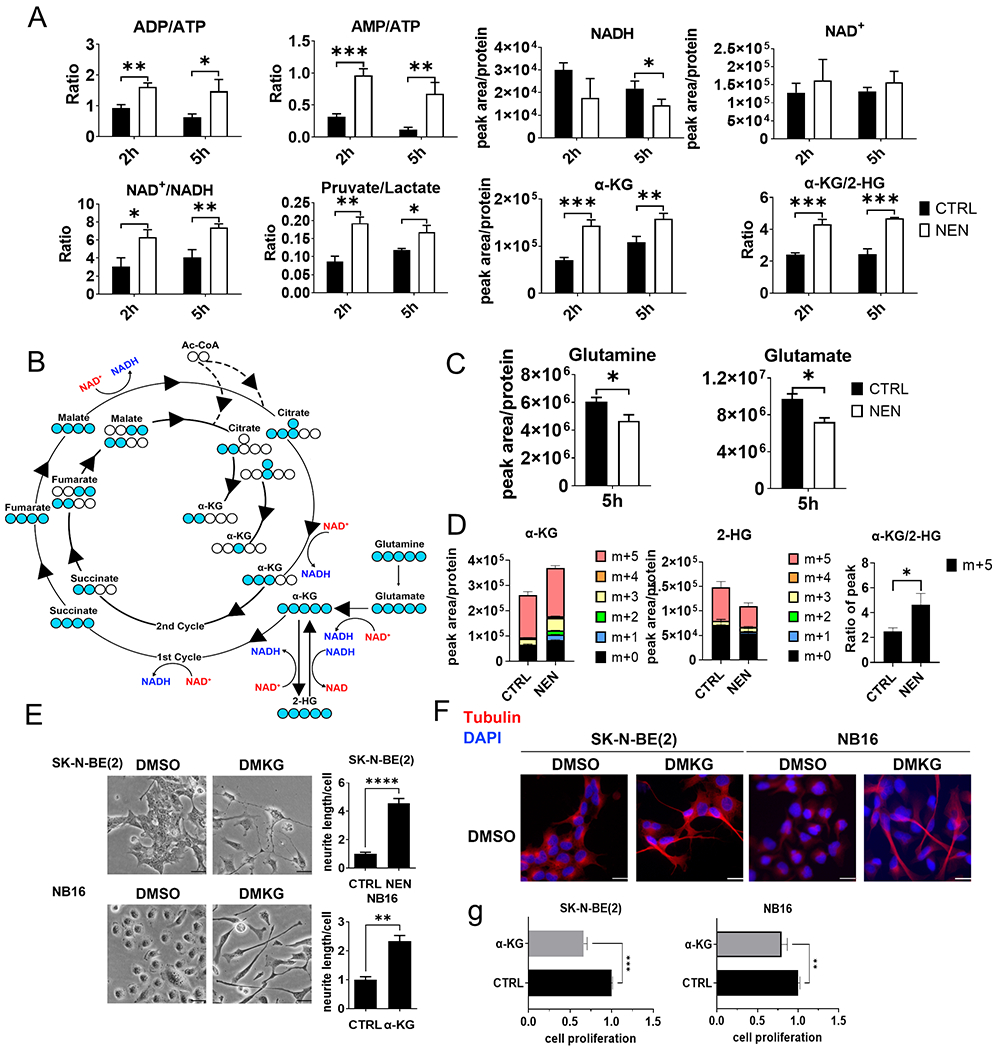
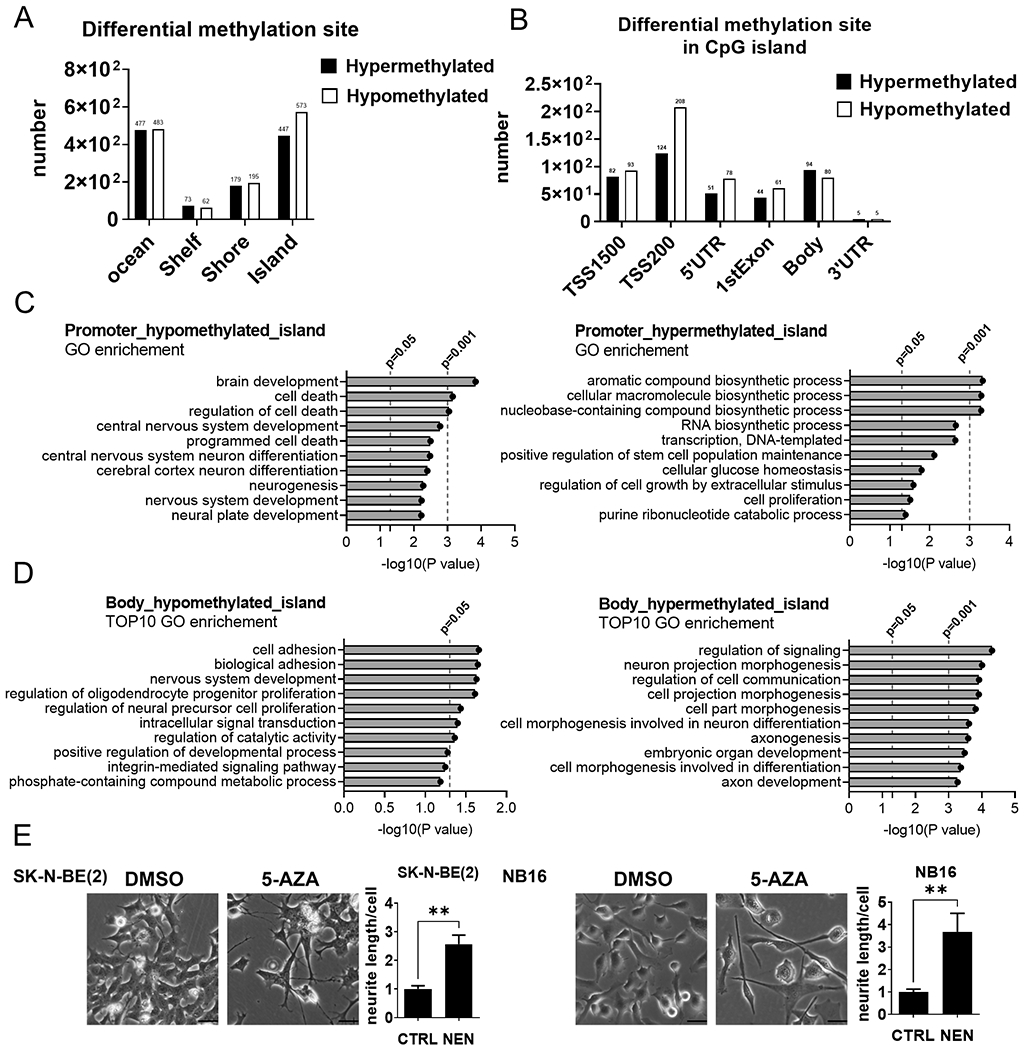
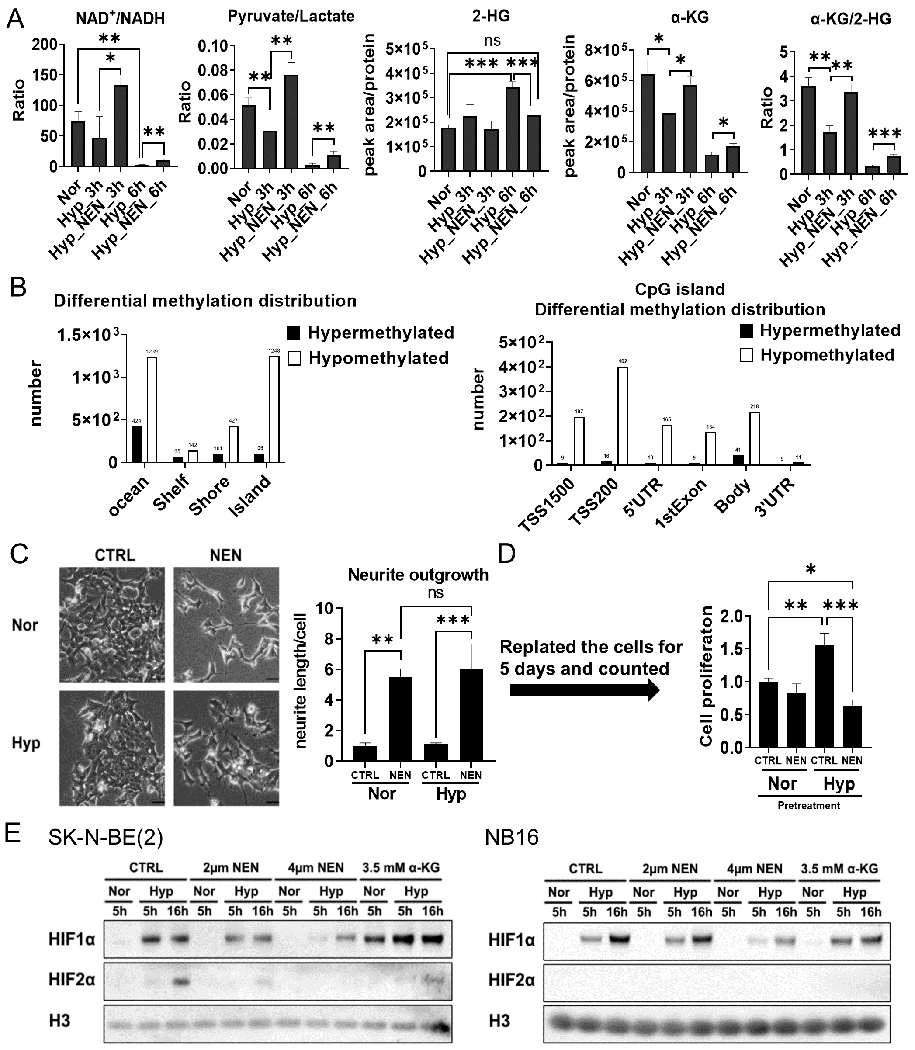
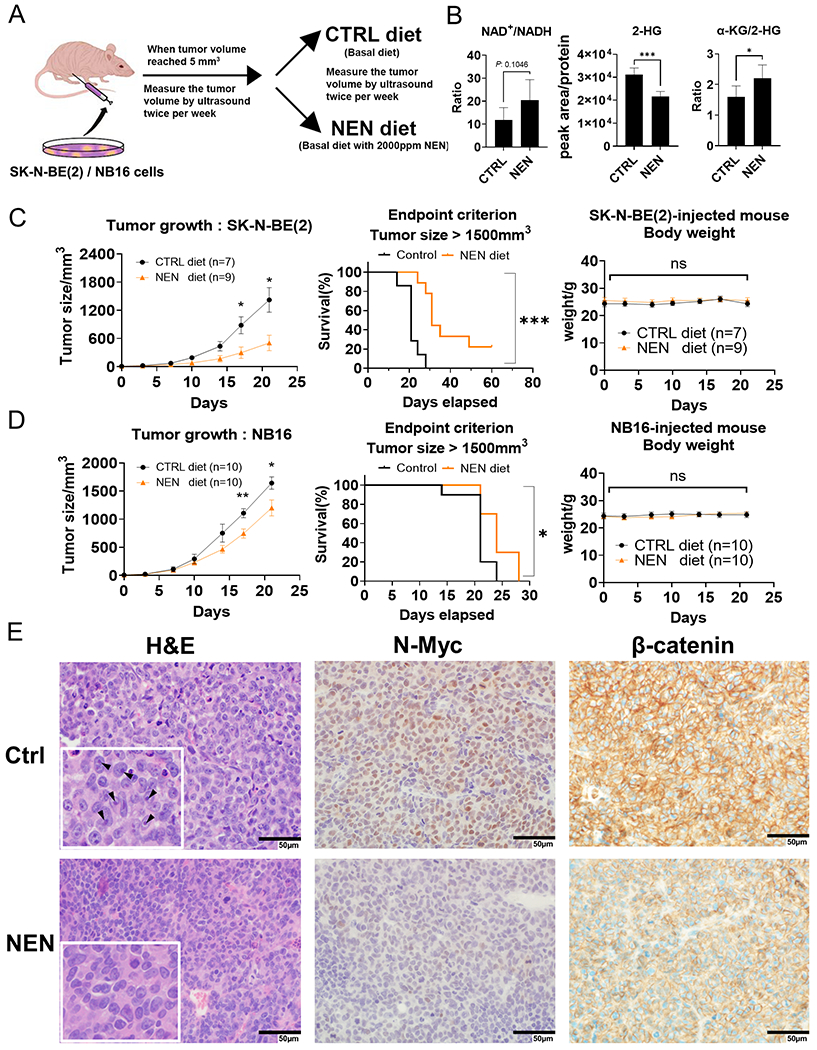
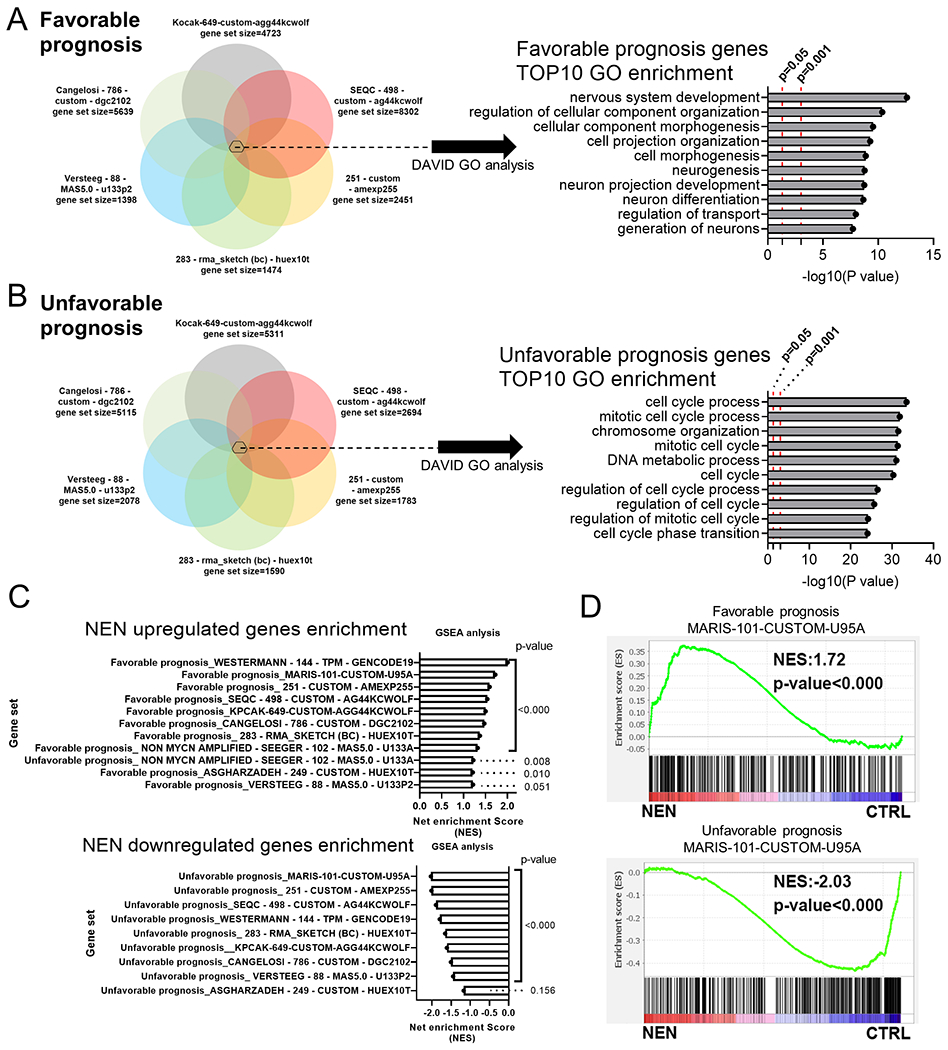
The Warburg effect is the major metabolic hallmark of cancer. According to Warburg himself, the consequence of the Warburg effect is cell dedifferentiation. Therefore, reversing the Warburg effect might be an approach to restore cell differentiation in cancer. In this study, we used a mitochondrial uncoupler, niclosamide ethanolamine (NEN), to activate mitochondrial respiration, which induced neural differentiation in neuroblastoma cells. NEN treatment increased the NAD+/NADH and pyruvate/lactate ratios and also the α-ketoglutarate/2-hydroxyglutarate (2-HG) ratio. Consequently, NEN treatment induced promoter CpG island demethylation and epigenetic landscape remodeling, activating the neural differentiation program. In addition, NEN treatment upregulated p53 but downregulated N-Myc and β-catenin signaling in neuroblastoma cells. Importantly, even under hypoxia, NEN treatment remained effective in inhibiting 2-HG generation, promoting DNA demethylation, and suppressing hypoxia-inducible factor signaling. Dietary NEN intervention reduced tumor growth rate, 2-HG levels, and expression of N-Myc and β-catenin in tumors in an orthotopic neuroblastoma mouse model. Integrative analysis indicated that NEN treatment upregulated favorable prognosis genes and downregulated unfavorable prognosis genes, which were defined using multiple neuroblastoma patient datasets. Altogether, these results suggest that mitochondrial uncoupling is an effective metabolic and epigenetic therapy for reversing the Warburg effect and inducing differentiation in neuroblastoma.
Significance: Targeting cancer metabolism using the mitochondrial uncoupler niclosamide ethanolamine leads to methylome reprogramming and differentiation in neuroblastoma, providing a therapeutic opportunity to reverse the Warburg effect and suppress tumor growth. See related commentary by Byrne and Bell, p.167.
©2022 American Association for Cancer Research.
Conflict of interest statement
Competing interests
H.J., Y.L. and J.Y. submitted a patent application related to this manuscript.
Figures
Comment in
-
Neuroblastoma Differentiation: The Untapped Potential of Mitochondrial Uncouplers.Cancer Res. 2023 Jan 18;83(2):167-169. doi: 10.1158/0008-5472.CAN-22-3350. Cancer Res. 2023. PMID: 36651076
References
-
- Warburg O, Posener K, Negelein E. On the metabolism of carcinoma cells. Biochem Z 1924;152:309–44
-
- Warburg O. On the origin of cancer cells. Science 1956;123:309–14 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
