

Untangling the roles of TOP2A and TOP2B in transcription and cancer
- PMID: 36322662
- PMCID: PMC9629710
- DOI: 10.1126/sciadv.add4920
Untangling the roles of TOP2A and TOP2B in transcription and cancer
Abstract
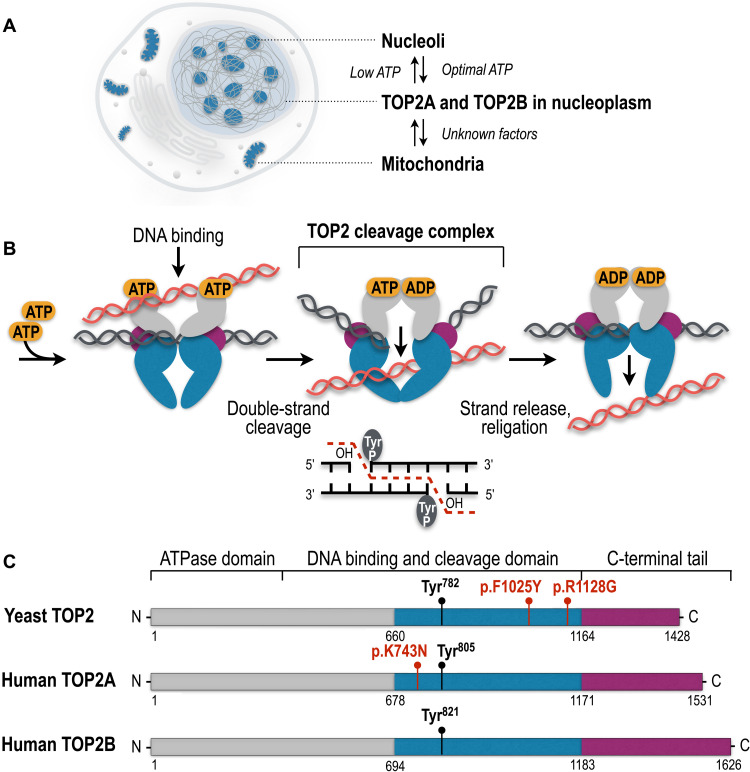
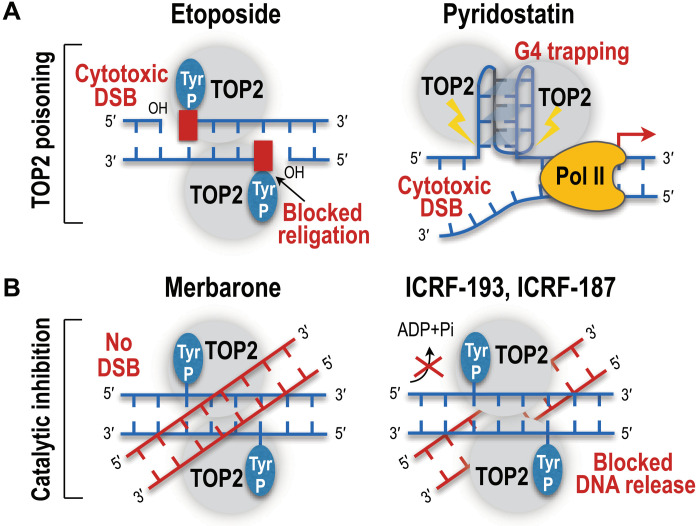
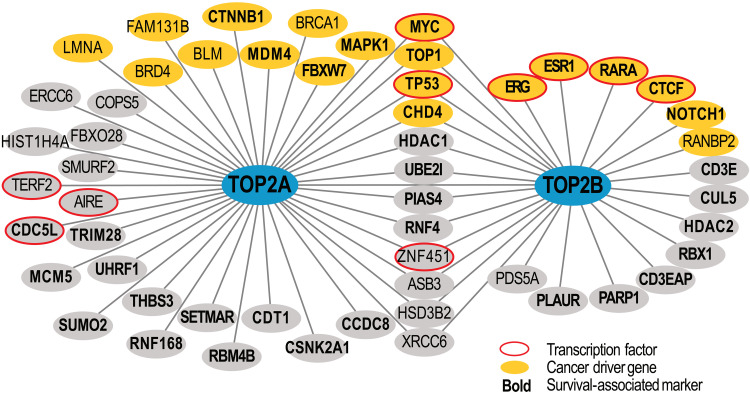
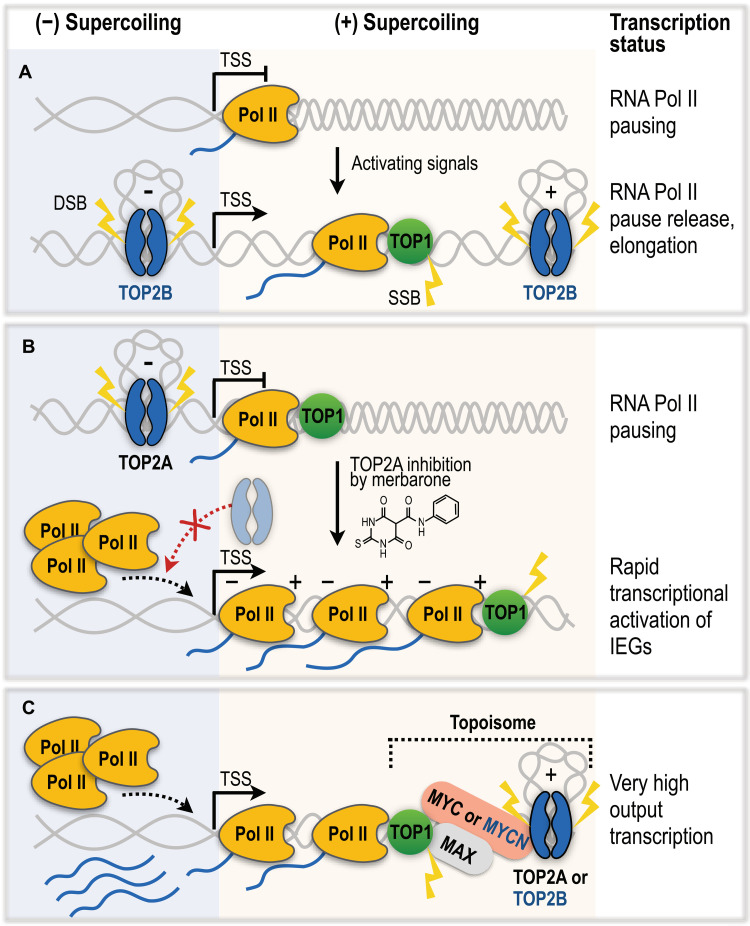
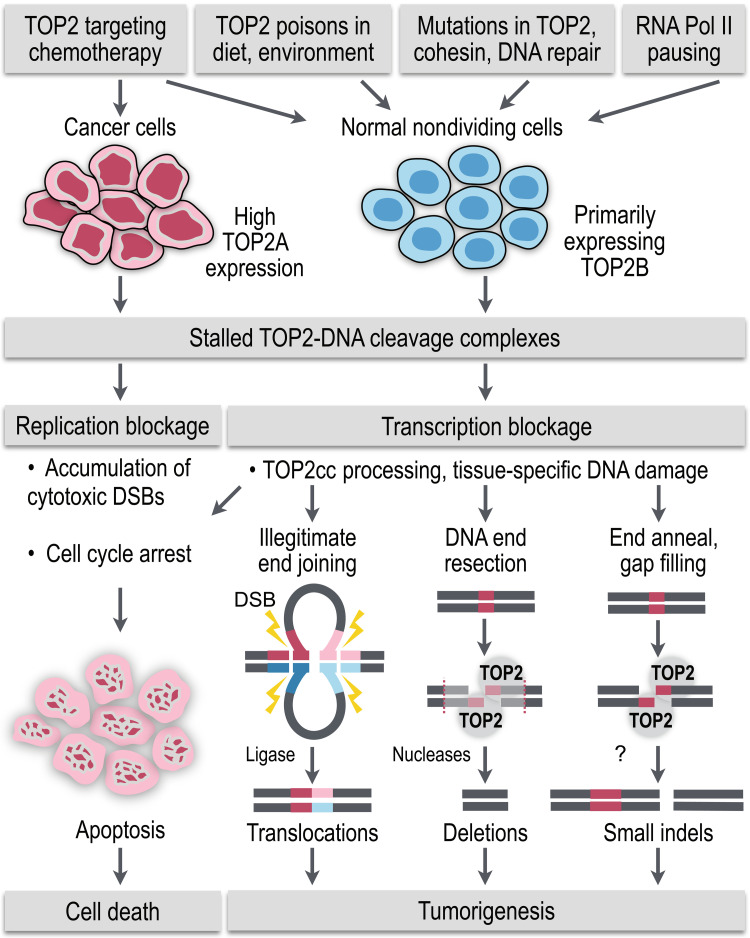
Type II topoisomerases (TOP2) are conserved regulators of chromatin topology that catalyze reversible DNA double-strand breaks (DSBs) and are essential for maintaining genomic integrity in diverse dynamic processes such as transcription, replication, and cell division. While controlled TOP2-mediated DSBs are an elegant solution to topological constraints of DNA, DSBs also contribute to the emergence of chromosomal translocations and mutations that drive cancer. The central importance of TOP2 enzymes as frontline chemotherapeutic targets is well known; however, their precise biological functions and impact in cancer development are still poorly understood. In this review, we provide an updated overview of TOP2A and TOP2B in the regulation of chromatin topology and transcription, and discuss the recent discoveries linking TOP2 activities with cancer pathogenesis.
Figures
References
-
- Champoux J. J., DNA topoisomerases: Structure, function, and mechanism. Annu. Rev. Biochem. 70, 369–413 (2001). - PubMed
-
- Ashour M. E., Atteya R., El-Khamisy S. F., Topoisomerase-mediated chromosomal break repair: An emerging player in many games. Nat. Rev. Cancer 15, 137–151 (2015). - PubMed
-
- Forterre P., Gribaldo S., Gadelle D., Serre M.-C., Origin and evolution of DNA topoisomerases. Biochimie 89, 427–446 (2007). - PubMed
-
- Corbett K. D., Berger J. M., Structure, molecular mechanisms, and evolutionary relationships in DNA topoisomerases. Annu. Rev. Biophys. Biomol. Struct. 33, 95–118 (2004). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
