

Quinacrine and Curcumin in combination decreased the breast cancer angiogenesis by modulating ABCG2 via VEGF A
- PMID: 36326988
- PMCID: PMC10409692
- DOI: 10.1007/s12079-022-00692-0
Quinacrine and Curcumin in combination decreased the breast cancer angiogenesis by modulating ABCG2 via VEGF A
Abstract
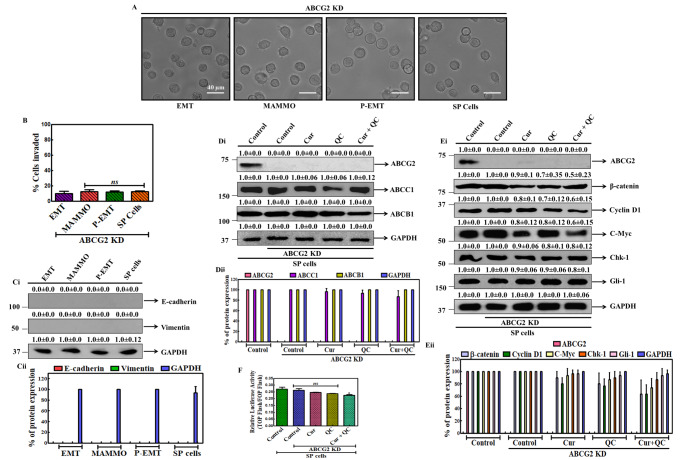
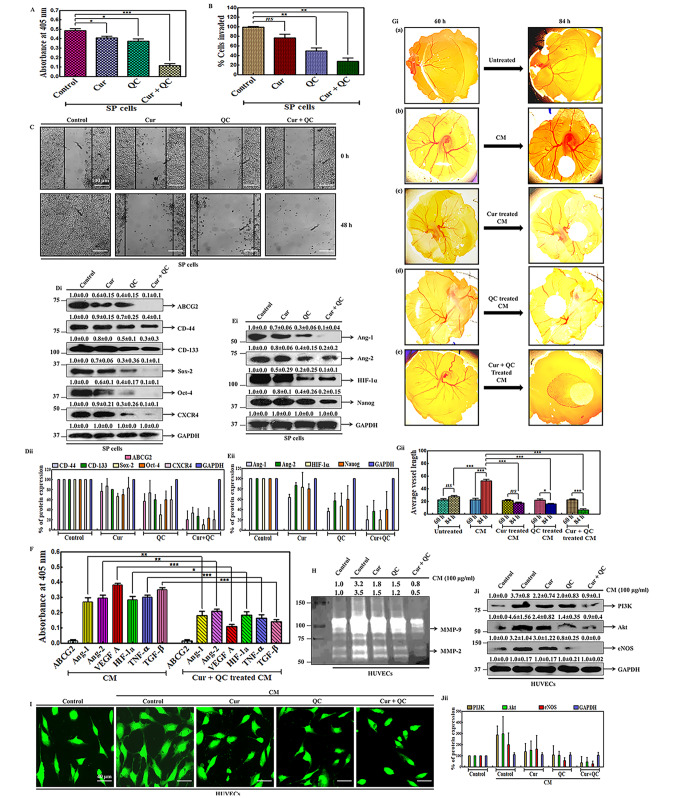
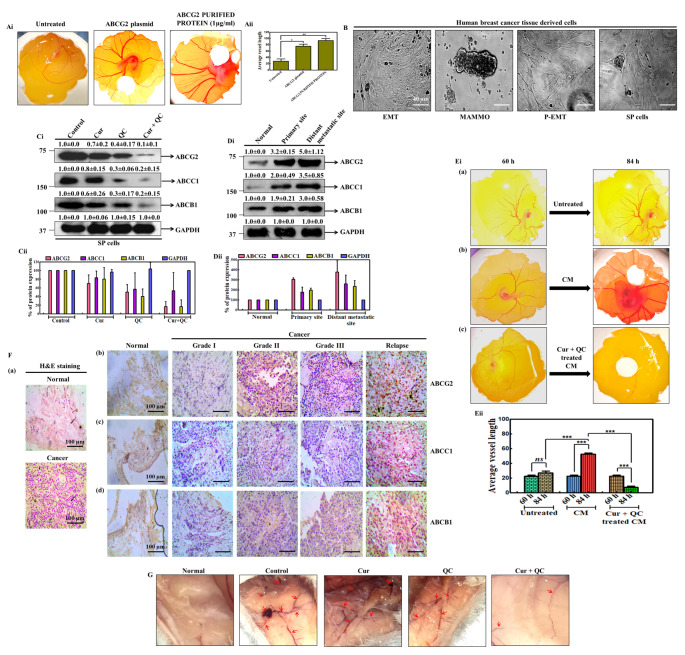
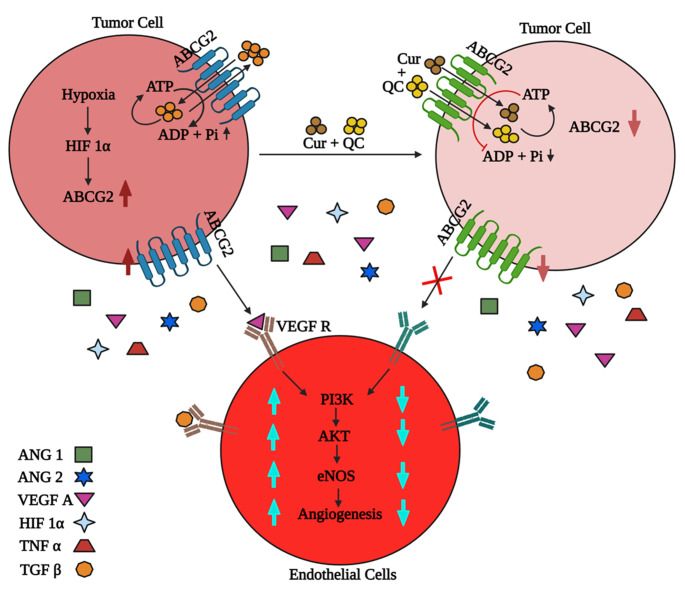
Cancer stem cells (CSCs) cause drug resistance in cancer due to its extensive drug efflux, DNA repair and self-renewal capability. ATP binding cassette subfamily G member 2 (ABCG2) efflux pump afford protection to CSCs in tumors, shielding them from the adverse effects of chemotherapy. Although the role of ABCG2 in cancer progression, invasiveness, recurrence are known but its role in metastasis and angiogenesis are not clear. Here, using in vitro (CSCs enriched side population [SP] cells), ex vivo (patient derived primary cells), in ovo (fertilized egg embryo) and in vivo (patient derived primary tissue mediated xenograft (PDX)) system, we have systematically studied the role of ABCG2 in angiogenesis and the regulation of the process by Curcumin (Cur) and Quinacrine (QC). Cur + QC inhibited the proliferation, invasion, migration and expression of representative markers of metastasis and angiogenesis. Following hypoxia, ABCG2 enriched cells released angiogenic factor vascular endothelial growth factor A (VEGF A) and induced the angiogenesis via PI3K-Akt-eNOS cascade. Cur + QC inhibited the ABCG2 expression and thus reduced the angiogenesis. Interestingly, overexpression of ABCG2 in SP cells and incubation of purified ABCG2 protein in media induced the angiogenesis but knockdown of ABCG2 decreased the vascularization. In agreement with in vitro results, ex vivo data showed similar phenomena. An induction of vascularization was noticed in PDX mice but reduction of vascularization was also observed after treatment of Cur + QC. Thus, data suggested that in hypoxia, ABCG2 enhances the production of angiogenesis factor VEGF A which in turn induced angiogenesis and Cur + QC inhibited the process by inhibiting ABCG2 in breast cancer.
Keywords: ABCG2; Angiogenesis; Cancer stem cells; Curcumin; HUVECs; Quinacrine.
© 2022. The International CCN Society.
Conflict of interest statement
The authors have no competing financial and/or non-financial interests.
Figures


References
-
- Chen Z, Liu F, Ren Q, et al. Suppression of ABCG2 inhibits cancer cell proliferation. Int J Cancer. 2010;126:841–851. - PubMed
-
- Chikazawa N, Tanaka H, Tasaka T, et al. Inhibition of Wnt signaling pathway decreases chemotherapy-resistant side-population colon cancer cells. Anticancer Res. 2010;30:2041–2048. - PubMed
LinkOut - more resources
Full Text Sources
