

MdbHLH4 negatively regulates apple cold tolerance by inhibiting MdCBF1/3 expression and promoting MdCAX3L-2 expression
- PMID: 36331333
- PMCID: PMC9806570
- DOI: 10.1093/plphys/kiac512
MdbHLH4 negatively regulates apple cold tolerance by inhibiting MdCBF1/3 expression and promoting MdCAX3L-2 expression
Abstract
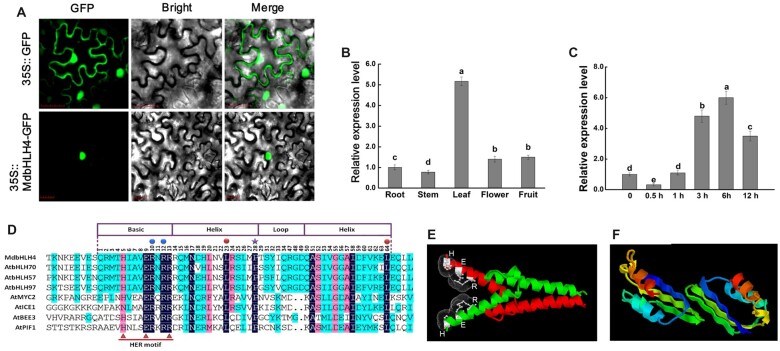
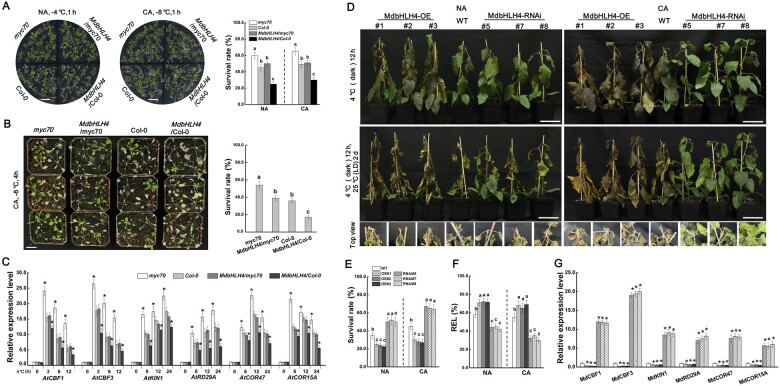
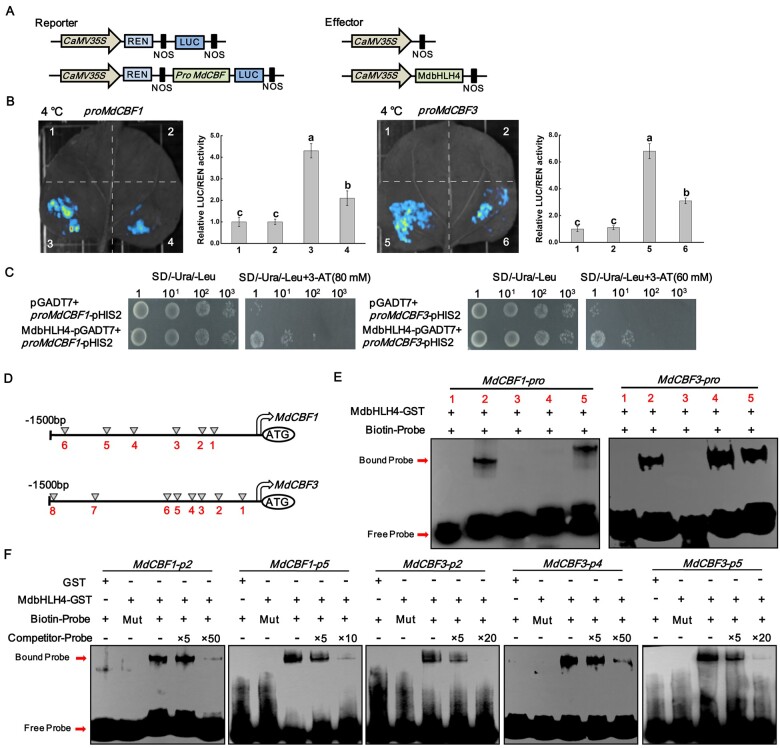
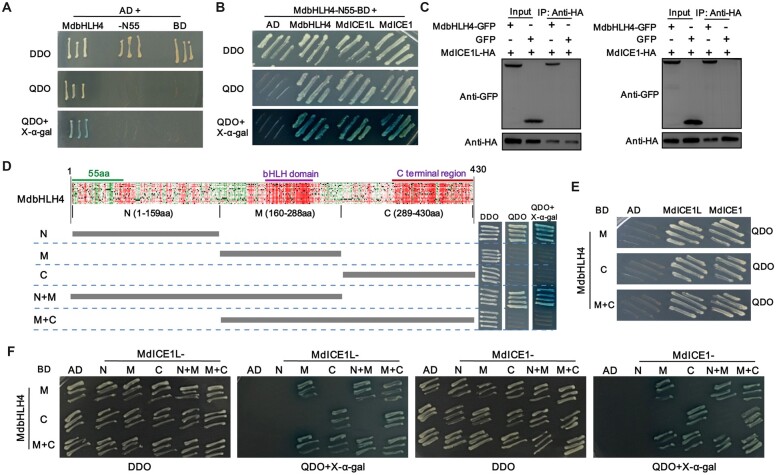
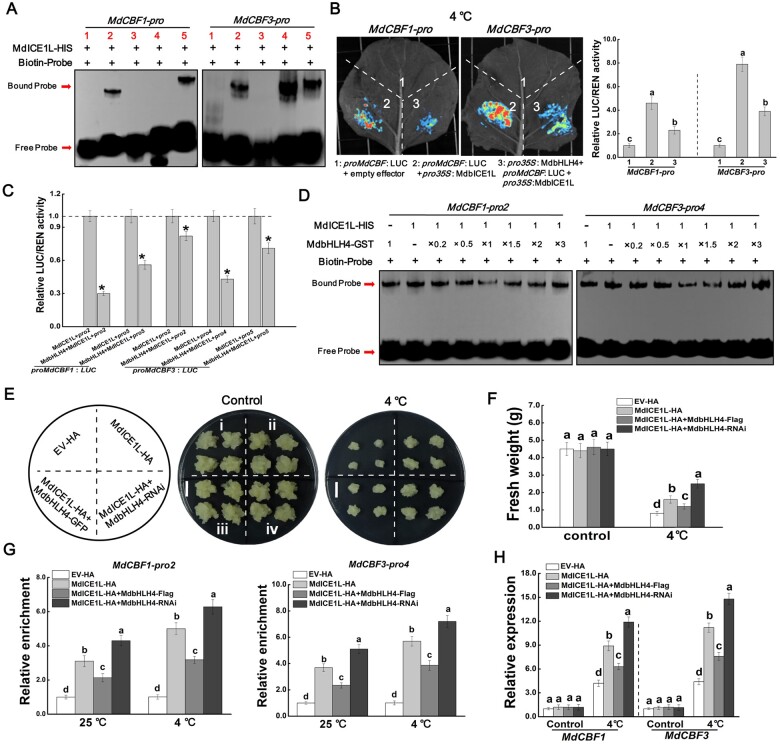
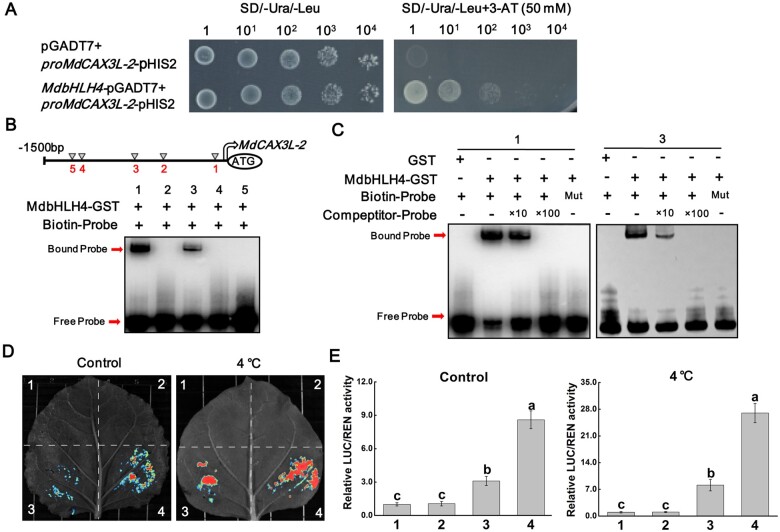
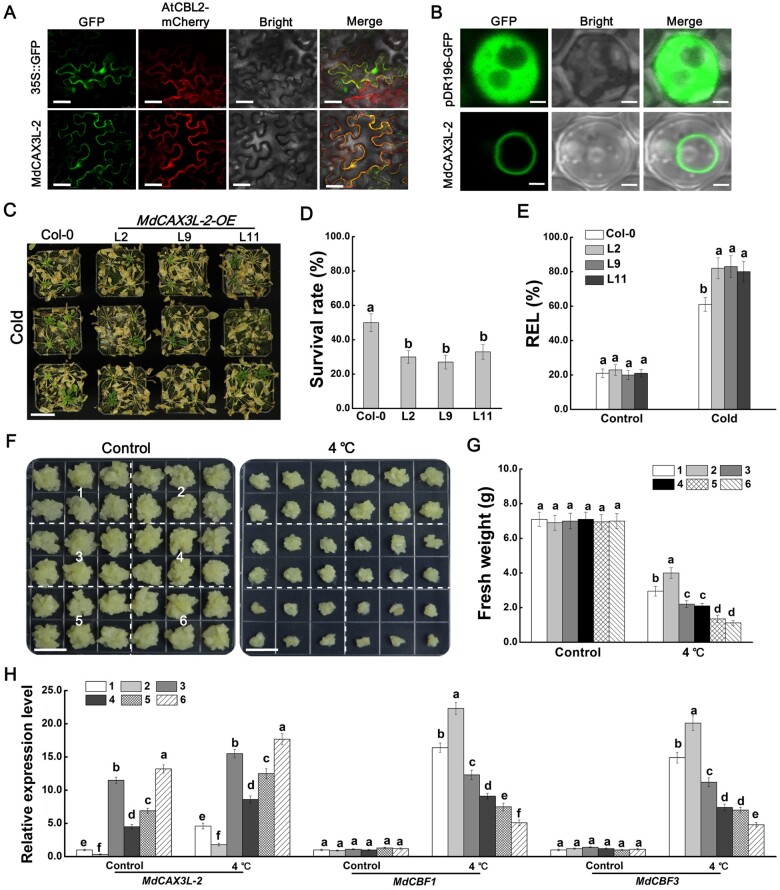
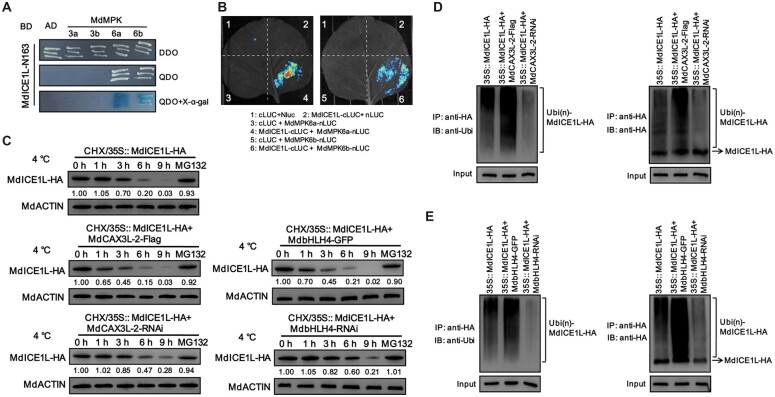
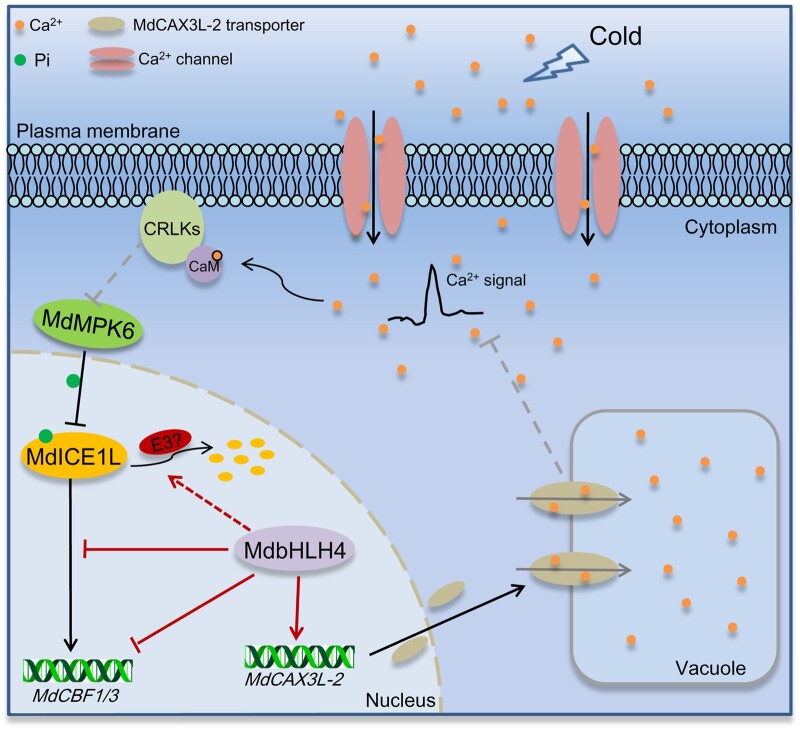
Low temperature affects the yield and quality of crops. Inducer of CBF expression 1 (ICE1) plays a positive role in plant cold tolerance by promoting the expression of CRT binding factor (CBF) and cold-responsive (COR) genes. Several ICE1-interacting transcription factors (TFs) that regulate plant cold tolerance have been identified. However, how these TFs affect the function of ICE1 and CBF expression under cold conditions remains unclear. Here, we identified the MYC-type TF MdbHLH4, a negative regulator of cold tolerance in Arabidopsis (Arabidopsis thaliana) and apple (Malus domestica) plants. Under cold conditions, MdbHLH4 inhibits the expression of MdCBF1 and MdCBF3 by directly binding to their promoters. It also interacts with MdICE1L, a homolog of AtICE1 in apple, and inhibits the binding of MdICE1L to the promoters of MdCBF1/3 and thus their expression. We showed that MdCAX3L-2, a Ca2+/H+ exchanger (CAX) family gene that negatively regulates plant cold tolerance, is also a direct target of MdbHLH4. MdbHLH4 reduced apple cold tolerance by promoting MdCAX3L-2 expression. Moreover, overexpression of either MdCAX3L-2 or MdbHLH4 promoted the cold-induced ubiquitination and degradation of MdICE1L. Overall, our results reveal that MdbHLH4 negatively regulates plant cold tolerance by inhibiting MdCBF1/3 expression and MdICE1L promoter-binding activity, as well as by promoting MdCAX3L-2 expression and cold-induced MdICE1L degradation. These findings provide insights into the mechanisms by which ICE1-interacting TFs regulate CBF expression and ICE1 function and thus plant cold tolerance.
© American Society of Plant Biologists 2022. All rights reserved. For permissions, please email: journals.permissions@oup.com.
Figures
Similar articles
-
The MdHB7L-MdICE1L-MdHOS1 Module Fine-Tunes Apple Cold Response via CBF-Dependent and CBF-Independent Pathways.Adv Sci (Weinh). 2025 Jul;12(25):e2501524. doi: 10.1002/advs.202501524. Epub 2025 Apr 26. Adv Sci (Weinh). 2025. PMID: 40285577 Free PMC article.
-
Apple B-box protein BBX37 regulates jasmonic acid mediated cold tolerance through the JAZ-BBX37-ICE1-CBF pathway and undergoes MIEL1-mediated ubiquitination and degradation.New Phytol. 2021 Mar;229(5):2707-2729. doi: 10.1111/nph.17050. Epub 2020 Dec 12. New Phytol. 2021. PMID: 33119890
-
An apple NAC transcription factor negatively regulates cold tolerance via CBF-dependent pathway.J Plant Physiol. 2018 Feb;221:74-80. doi: 10.1016/j.jplph.2017.12.009. Epub 2017 Dec 13. J Plant Physiol. 2018. PMID: 29253732
-
CBF-dependent signaling pathway: a key responder to low temperature stress in plants.Crit Rev Biotechnol. 2011 Jun;31(2):186-92. doi: 10.3109/07388551.2010.505910. Epub 2010 Oct 4. Crit Rev Biotechnol. 2011. PMID: 20919819 Review.
-
Cold-responsive gene regulation during cold acclimation in plants.Plant Signal Behav. 2010 Aug;5(8):948-52. doi: 10.4161/psb.5.8.12135. Epub 2010 Aug 1. Plant Signal Behav. 2010. PMID: 20699657 Free PMC article. Review.
Cited by
-
Melatonin enhances KCl salinity tolerance by maintaining K+ homeostasis in Malus hupehensis.Plant Biotechnol J. 2023 Nov;21(11):2273-2290. doi: 10.1111/pbi.14129. Epub 2023 Jul 19. Plant Biotechnol J. 2023. PMID: 37465981 Free PMC article.
-
Identification of MYC genes in four Cucurbitaceae species and their roles in the response to temperature stress.BMC Genomics. 2024 Sep 16;25(1):867. doi: 10.1186/s12864-024-10771-8. BMC Genomics. 2024. PMID: 39285374 Free PMC article.
-
MdNAC104 positively regulates apple cold tolerance via CBF-dependent and CBF-independent pathways.Plant Biotechnol J. 2023 Oct;21(10):2057-2073. doi: 10.1111/pbi.14112. Epub 2023 Jun 30. Plant Biotechnol J. 2023. PMID: 37387580 Free PMC article.
-
EjFAD8 Enhances the Low-Temperature Tolerance of Loquat by Desaturation of Sulfoquinovosyl Diacylglycerol (SQDG).Int J Mol Sci. 2023 Apr 8;24(8):6946. doi: 10.3390/ijms24086946. Int J Mol Sci. 2023. PMID: 37108110 Free PMC article.
-
Ectopic expression of HvbHLH132 from hulless barley reduces cold tolerance in transgenic Arabidopsis thaliana.Plant Cell Rep. 2024 Nov 25;43(12):297. doi: 10.1007/s00299-024-03382-w. Plant Cell Rep. 2024. PMID: 39585367
References
-
- Agarwal M, Hao YJ, Kapoor A, Dong CH, Fujii H, Zheng XW, Zhu JK (2006) A R2R3 type MYB transcription factor is involved in the cold regulation of CBF genes and in acquired freezing tolerance. J Biol Chem 281: 37636–37645 - PubMed
-
- An JP, Wang XF, Zhang XW, You CX, Hao YJ (2021) Apple B-box protein BBX37 regulates jasmonic acid mediated cold tolerance through the JAZ-BBX37-ICE1-CBF pathway and undergoes MIEL1-mediated ubiquitination and degradation. New Phytol 229: 2707–2729 - PubMed
-
- Chen XX, Ding YL, Yang YQ, Song CP, Wang BS, Yang SH, Guo Y, Gong ZZ (2021) Protein kinases in plant responses to drought, salt, and cold stress. J Integr Plant Biol 63: 53–78 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
