

Oxidized thioredoxin-1 restrains the NLRP1 inflammasome
- PMID: 36332009
- PMCID: PMC9850498
- DOI: 10.1126/sciimmunol.abm7200
Oxidized thioredoxin-1 restrains the NLRP1 inflammasome
Abstract
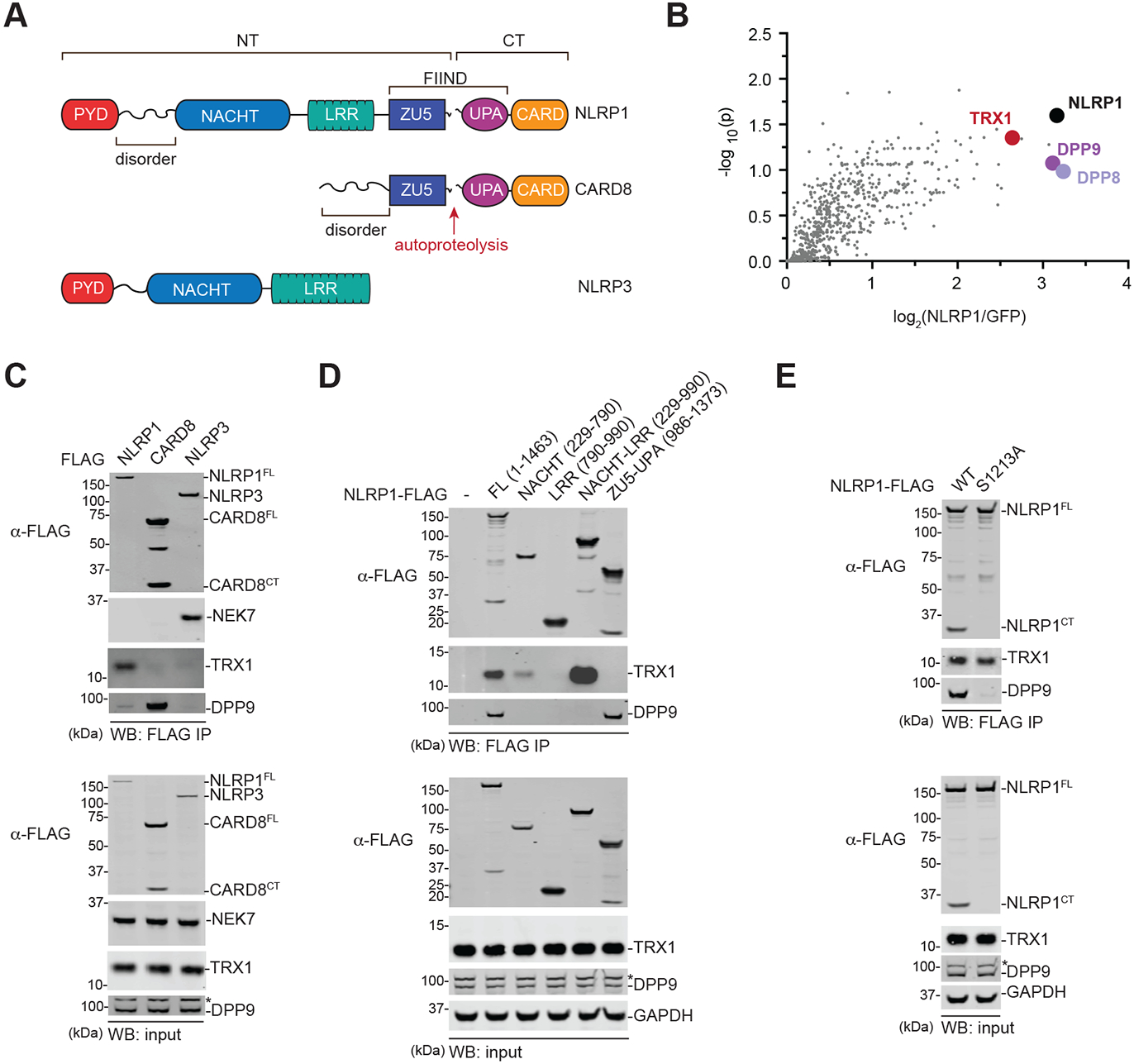
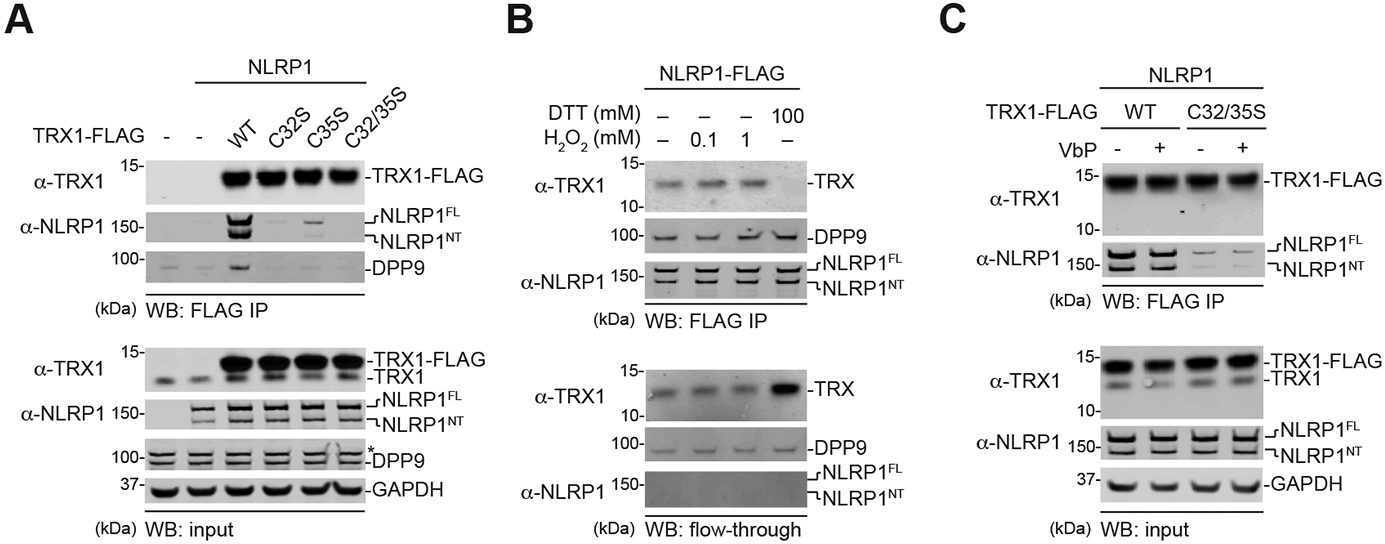
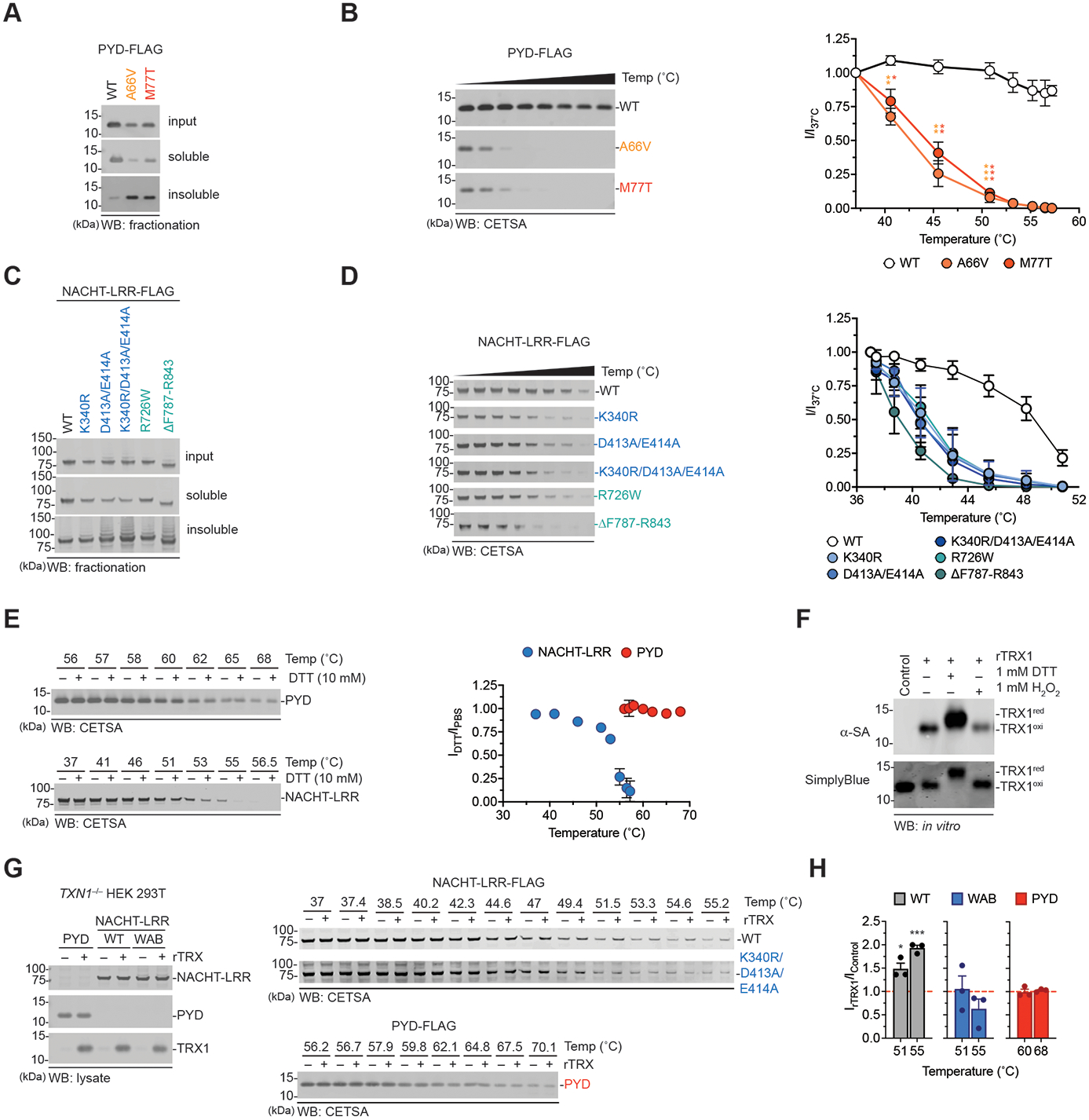
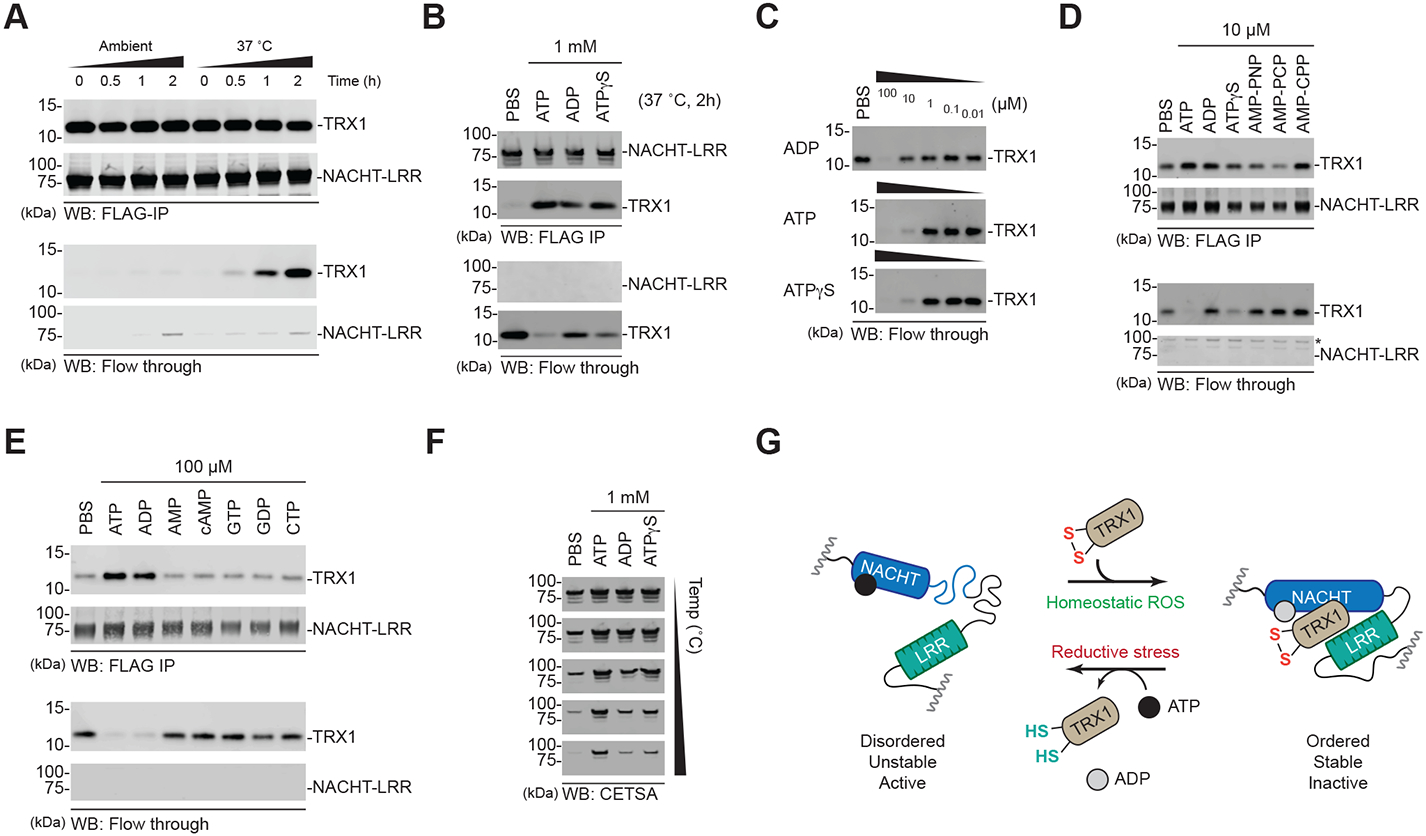
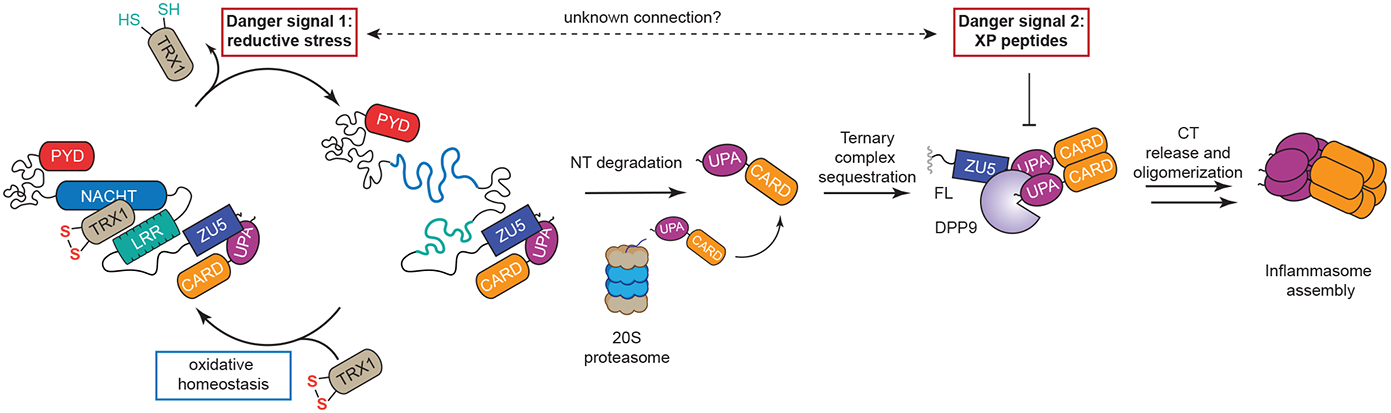
The danger signals that activate the NLRP1 inflammasome have not been established. Here, we report that the oxidized, but not the reduced, form of thioredoxin-1 (TRX1) binds to NLRP1. We found that oxidized TRX1 associates with the NACHT-LRR region of NLRP1 in an ATP-dependent process, forming a stable complex that restrains inflammasome activation. Consistent with these findings, patient-derived and ATPase-inactivating mutations in the NACHT-LRR region that cause hyperactive inflammasome formation interfere with TRX1 binding. Overall, this work strongly suggests that reductive stress, the cellular perturbation that will eliminate oxidized TRX1 and abrogate the TRX1-NLRP1 interaction, is a danger signal that activates the NLRP1 inflammasome.
Figures


References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
