

Insights into the evolution of enzymatic specificity and catalysis: From Asgard archaea to human adenylate kinases
- PMID: 36332013
- PMCID: PMC9635829
- DOI: 10.1126/sciadv.abm4089
Insights into the evolution of enzymatic specificity and catalysis: From Asgard archaea to human adenylate kinases
Abstract
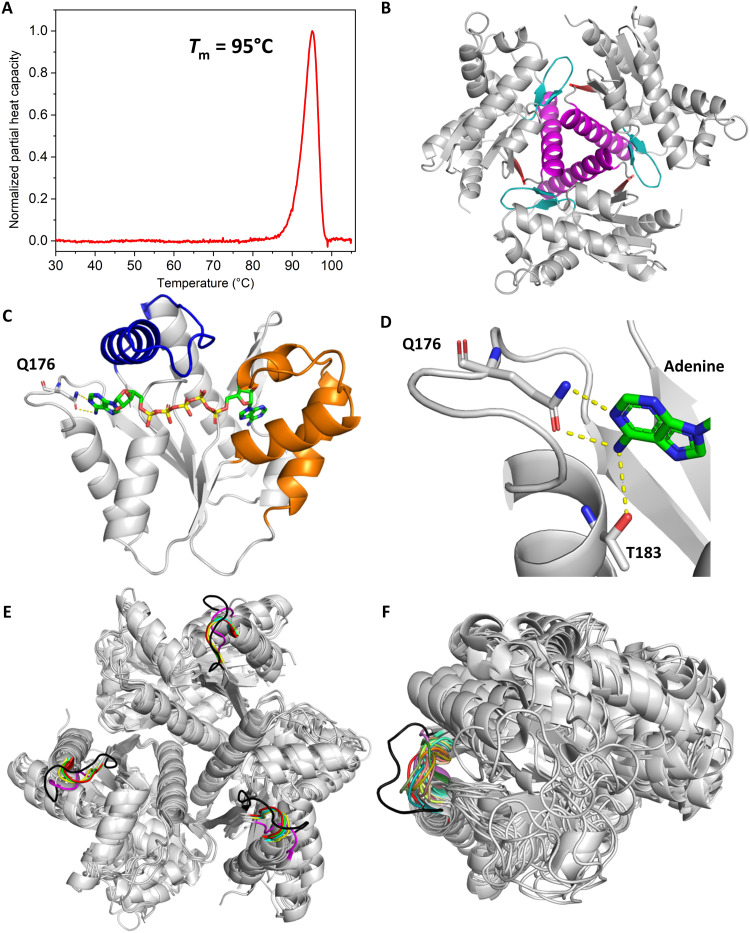
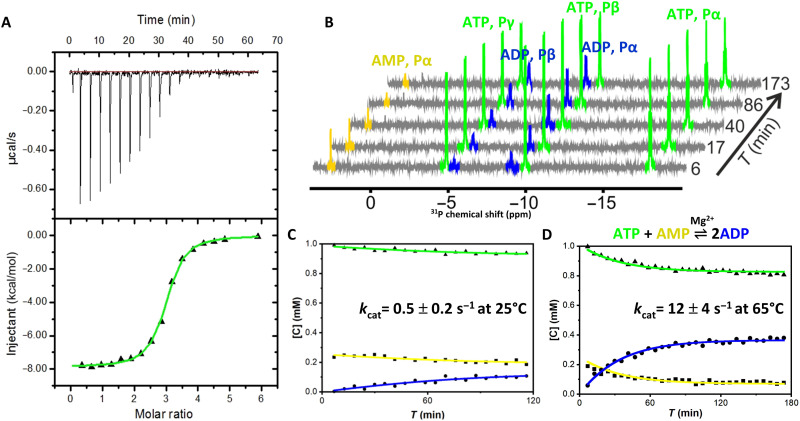
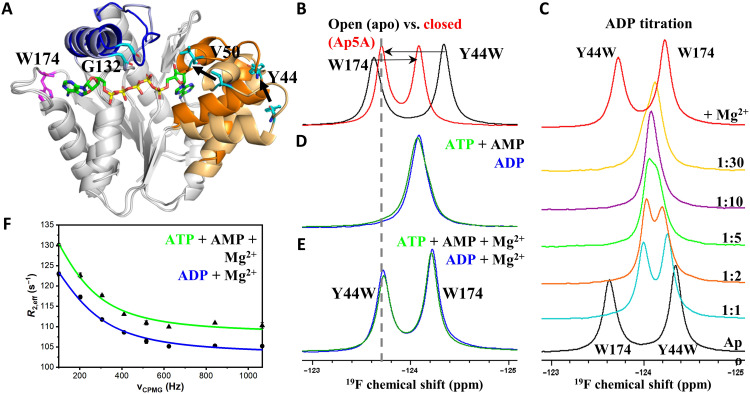
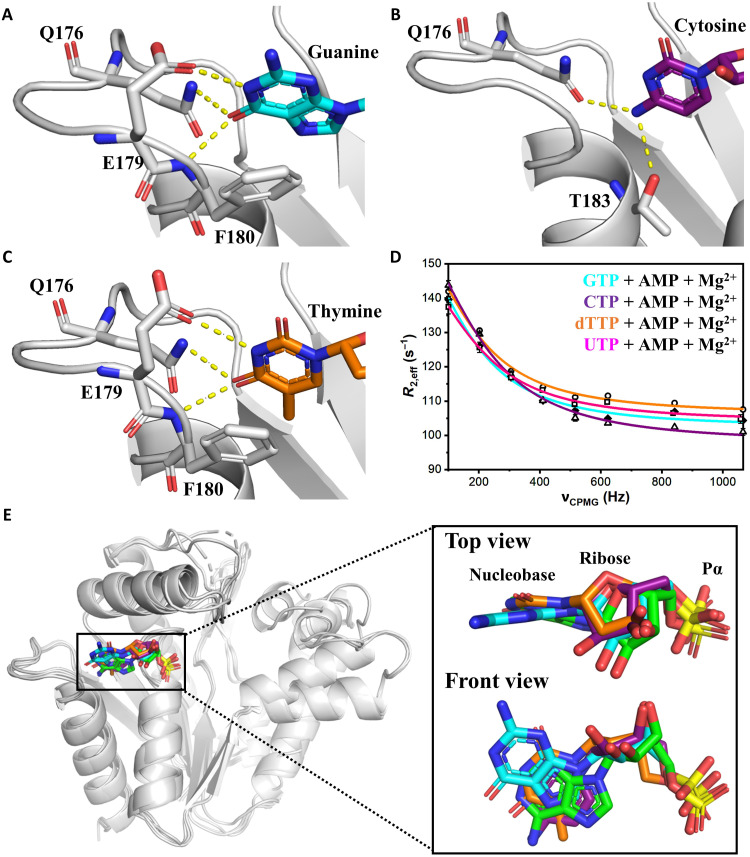
Enzymatic catalysis is critically dependent on selectivity, active site architecture, and dynamics. To contribute insights into the interplay of these properties, we established an approach with NMR, crystallography, and MD simulations focused on the ubiquitous phosphotransferase adenylate kinase (AK) isolated from Odinarchaeota (OdinAK). Odinarchaeota belongs to the Asgard archaeal phylum that is believed to be the closest known ancestor to eukaryotes. We show that OdinAK is a hyperthermophilic trimer that, contrary to other AK family members, can use all NTPs for its phosphorylation reaction. Crystallographic structures of OdinAK-NTP complexes revealed a universal NTP-binding motif, while 19F NMR experiments uncovered a conserved and rate-limiting dynamic signature. As a consequence of trimerization, the active site of OdinAK was found to be lacking a critical catalytic residue and is therefore considered to be "atypical." On the basis of discovered relationships with human monomeric homologs, our findings are discussed in terms of evolution of enzymatic substrate specificity and cold adaptation.
Figures

Similar articles
-
Diversity, origin, and evolution of the ESCRT systems.mBio. 2024 Mar 13;15(3):e0033524. doi: 10.1128/mbio.00335-24. Epub 2024 Feb 21. mBio. 2024. PMID: 38380930 Free PMC article.
-
"Sifarchaeota," a Novel Asgard Phylum from Costa Rican Sediment Capable of Polysaccharide Degradation and Anaerobic Methylotrophy.Appl Environ Microbiol. 2021 Apr 13;87(9):e02584-20. doi: 10.1128/AEM.02584-20. Print 2021 Apr 13. Appl Environ Microbiol. 2021. PMID: 33608286 Free PMC article.
-
Expanding Asgard members in the domain of Archaea sheds new light on the origin of eukaryotes.Sci China Life Sci. 2022 Apr;65(4):818-829. doi: 10.1007/s11427-021-1969-6. Epub 2021 Aug 6. Sci China Life Sci. 2022. PMID: 34378142
-
Asgard archaea: Diversity, function, and evolutionary implications in a range of microbiomes.AIMS Microbiol. 2019 Jan 30;5(1):48-61. doi: 10.3934/microbiol.2019.1.48. eCollection 2019. AIMS Microbiol. 2019. PMID: 31384702 Free PMC article. Review.
-
Structural Insights into the Molecular Evolution of the Archaeal Exo-β-d-Glucosaminidase.Int J Mol Sci. 2019 May 18;20(10):2460. doi: 10.3390/ijms20102460. Int J Mol Sci. 2019. PMID: 31109049 Free PMC article. Review.
Cited by
-
Perspectives on Computational Enzyme Modeling: From Mechanisms to Design and Drug Development.ACS Omega. 2024 Feb 8;9(7):7393-7412. doi: 10.1021/acsomega.3c09084. eCollection 2024 Feb 20. ACS Omega. 2024. PMID: 38405524 Free PMC article. Review.
-
Recent Advances in Biosensors Using Enzyme-Stabilized Gold Nanoclusters.Biosensors (Basel). 2024 Dec 24;15(1):2. doi: 10.3390/bios15010002. Biosensors (Basel). 2024. PMID: 39852053 Free PMC article. Review.
-
Magnesium induced structural reorganization in the active site of adenylate kinase.Sci Adv. 2024 Aug 9;10(32):eado5504. doi: 10.1126/sciadv.ado5504. Epub 2024 Aug 9. Sci Adv. 2024. PMID: 39121211 Free PMC article.
-
Protein dynamics: The future is bright and complicated!Struct Dyn. 2023 Feb 27;10(1):014301. doi: 10.1063/4.0000179. eCollection 2023 Jan. Struct Dyn. 2023. PMID: 36865927 Free PMC article.
-
Insights into Enzymatic Catalysis from Binding and Hydrolysis of Diadenosine Tetraphosphate by E. coli Adenylate Kinase.Biochemistry. 2023 Aug 1;62(15):2238-2243. doi: 10.1021/acs.biochem.3c00189. Epub 2023 Jul 7. Biochemistry. 2023. PMID: 37418448 Free PMC article.
References
-
- Rona P. A., Klinkhammer G., Nelsen T. A., Trefry J. H., Elderfield H., Black smokers, massive sulfides and vent biota at the Mid-Atlantic Ridge. Nature 321, 33–37 (1986).
-
- Zaremba-Niedzwiedzka K., Caceres E. F., Saw J. H., Bäckström D., Juzokaite L., Vancaester E., Seitz K. W., Anantharaman K., Starnawski P., Kjeldsen K. U., Stott M. B., Nunoura T., Banfield J. F., Schramm A., Baker B. J., Spang A., Ettema T. J. G., Asgard archaea illuminate the origin of eukaryotic cellular complexity. Nature 541, 353–358 (2017). - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
