

Architecture of the outbred brown fat proteome defines regulators of metabolic physiology
- PMID: 36334589
- PMCID: PMC10040263
- DOI: 10.1016/j.cell.2022.10.003
Architecture of the outbred brown fat proteome defines regulators of metabolic physiology
Abstract
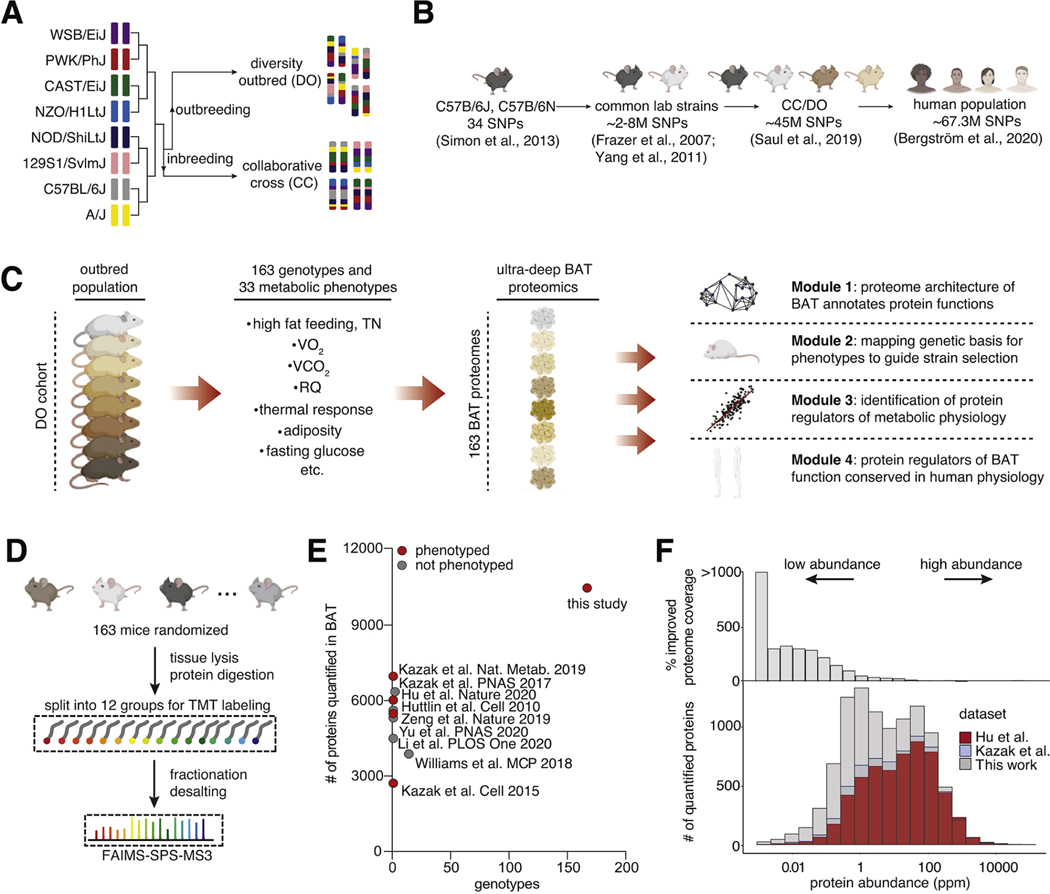
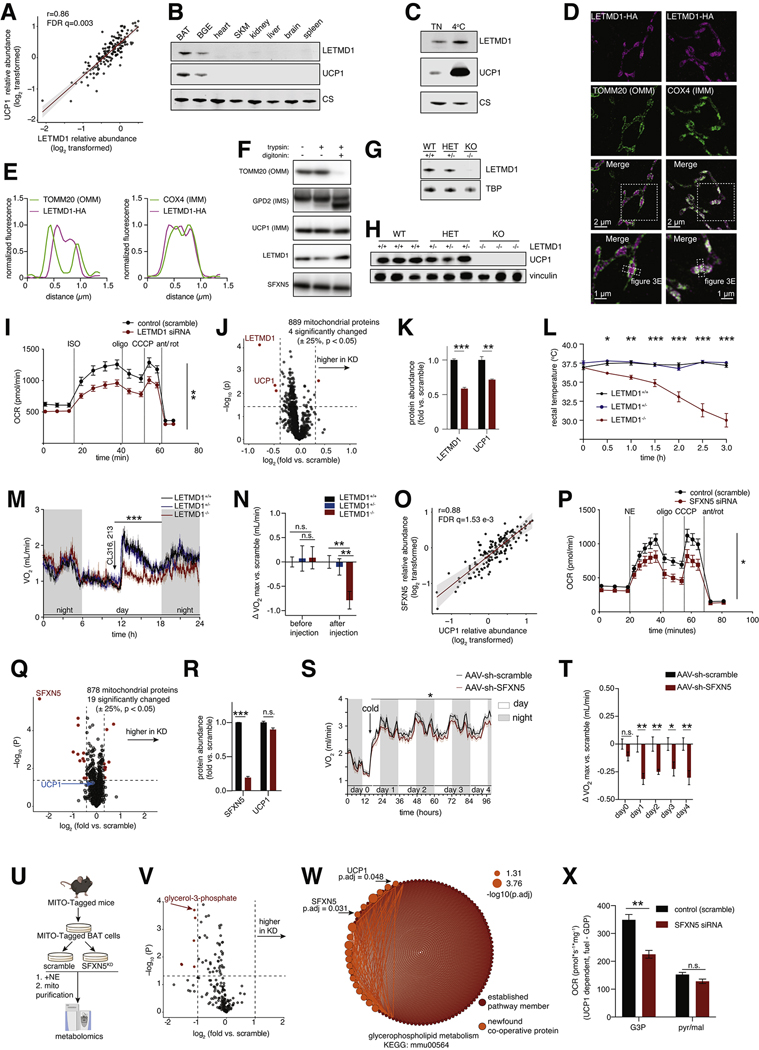
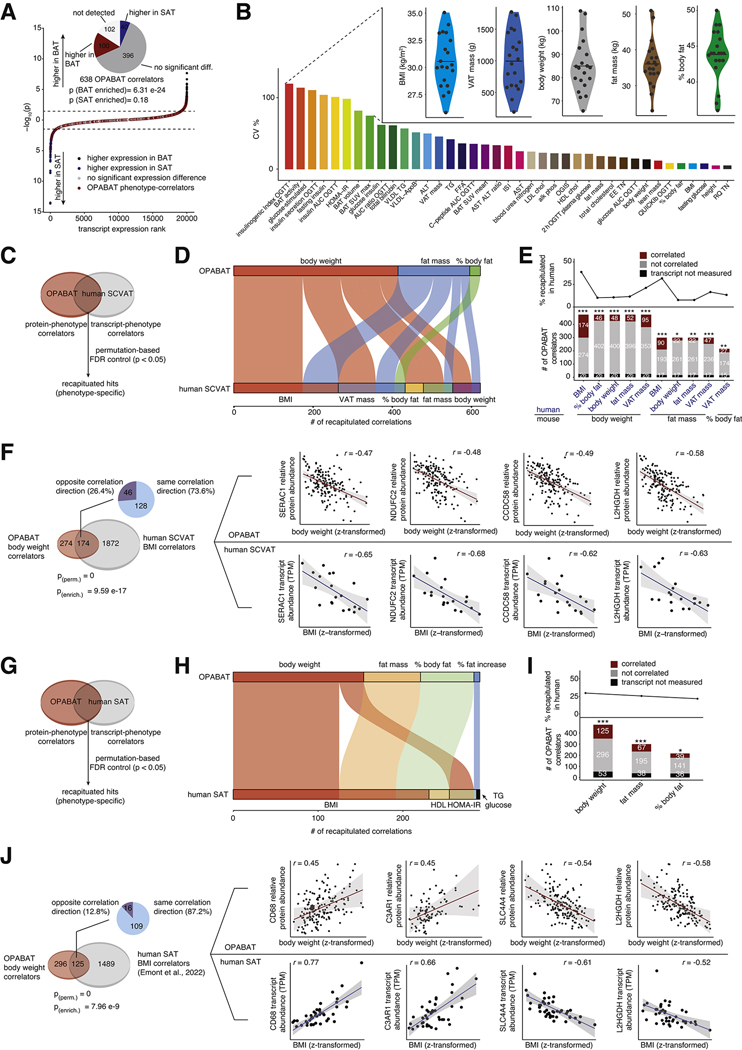
Brown adipose tissue (BAT) regulates metabolic physiology. However, nearly all mechanistic studies of BAT protein function occur in a single inbred mouse strain, which has limited the understanding of generalizable mechanisms of BAT regulation over physiology. Here, we perform deep quantitative proteomics of BAT across a cohort of 163 genetically defined diversity outbred mice, a model that parallels the genetic and phenotypic variation found in humans. We leverage this diversity to define the functional architecture of the outbred BAT proteome, comprising 10,479 proteins. We assign co-operative functions to 2,578 proteins, enabling systematic discovery of regulators of BAT. We also identify 638 proteins that correlate with protection from, or sensitivity to, at least one parameter of metabolic disease. We use these findings to uncover SFXN5, LETMD1, and ATP1A2 as modulators of BAT thermogenesis or adiposity, and provide OPABAT as a resource for understanding the conserved mechanisms of BAT regulation over metabolic physiology.
Copyright © 2022 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests F.E.M. and N.V.B. and are currently employees of Calico Life Sciences, LLC. T.B. is currently an employee of Roche Diagnostics. E.T.C. is a founder, equity holder, and consultant for Matchpoint Therapeutics and Aevum Therapeutics. B.M.S. is a founder, equity holder, and consultant for Aevum Therapeutics.
Figures




References
-
- Antonicka H, Lin ZY, Janer A, Aaltonen MJ, Weraarpachai W, Gingras AC, and Shoubridge EA (2020). A High-Density Human Mitochondrial Proximity Interaction Network. Cell Metab 32, 479–497 e479. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 DK123095/DK/NIDDK NIH HHS/United States
- U24 HG006673/HG/NHGRI NIH HHS/United States
- R01 DK102898/DK/NIDDK NIH HHS/United States
- P30 DK056341/DK/NIDDK NIH HHS/United States
- U41 HG006673/HG/NHGRI NIH HHS/United States
- RC2 DK116691/DK/NIDDK NIH HHS/United States
- P30 DK057521/DK/NIDDK NIH HHS/United States
- K99 AG073461/AG/NIA NIH HHS/United States
- P30 DK135043/DK/NIDDK NIH HHS/United States
- R01 DK087092/DK/NIDDK NIH HHS/United States
- P30 CA008748/CA/NCI NIH HHS/United States
- R01 DK102173/DK/NIDDK NIH HHS/United States
- P30 DK046200/DK/NIDDK NIH HHS/United States
- P30 DK036836/DK/NIDDK NIH HHS/United States
- K99 CA273170/CA/NCI NIH HHS/United States
- P30 DK098722/DK/NIDDK NIH HHS/United States
- T32 DK007516/DK/NIDDK NIH HHS/United States
- R01 GM070683/GM/NIGMS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
