

Brain molecular mechanisms in Rasmussen encephalitis
- PMID: 36336987
- PMCID: PMC9852002
- DOI: 10.1111/epi.17457
Brain molecular mechanisms in Rasmussen encephalitis
Abstract
Objective: This study was undertaken to identify molecular mechanisms in brain tissue of Rasmussen encephalitis (RE) when compared to people with non-RE epilepsy (PWE) and control cases using whole exome sequencing (WES), RNAseq, and proteomics.
Methods: Frozen brain tissue (ages = 2-19 years) was obtained from control autopsy (n = 14), surgical PWE (n = 10), and surgical RE cases (n = 27). We evaluated WES variants in RE associated with epilepsy, seizures, RE, and human leukocyte antigens (HLAs). Differential expression was evaluated by RNAseq (adjusted p < .05) and label-free quantitative mass spectrometry (false discovery rate < 5%) in the three groups.
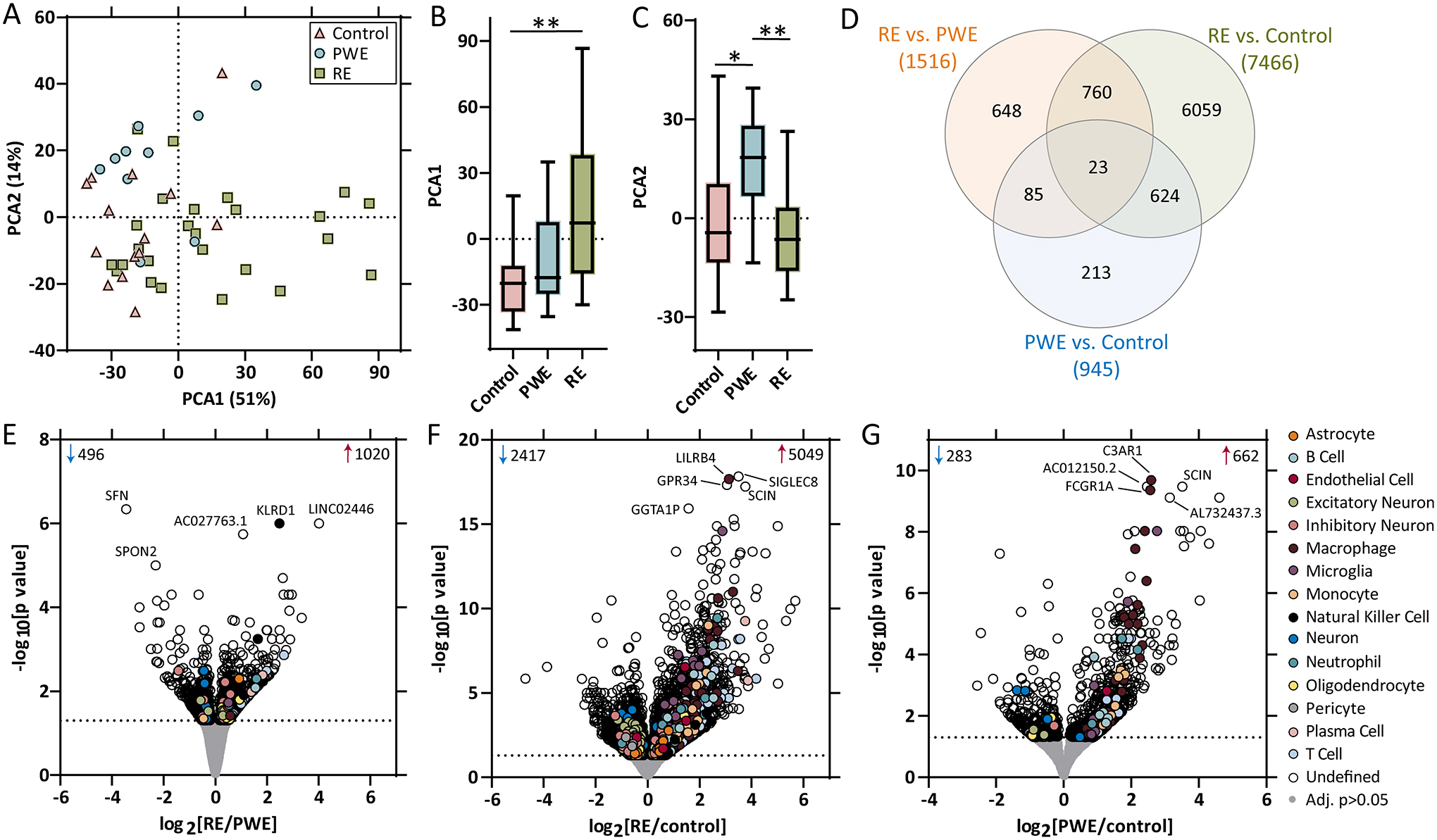
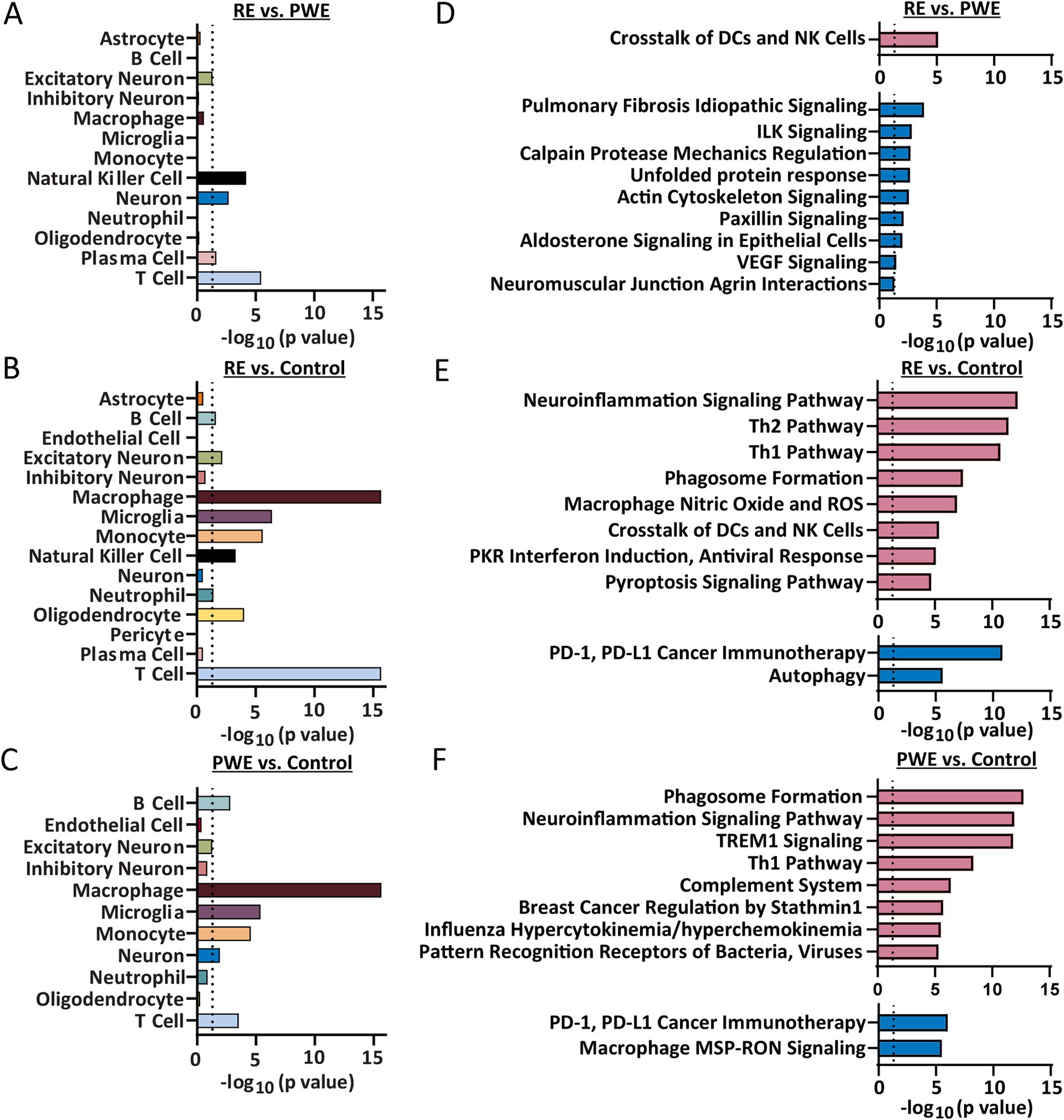
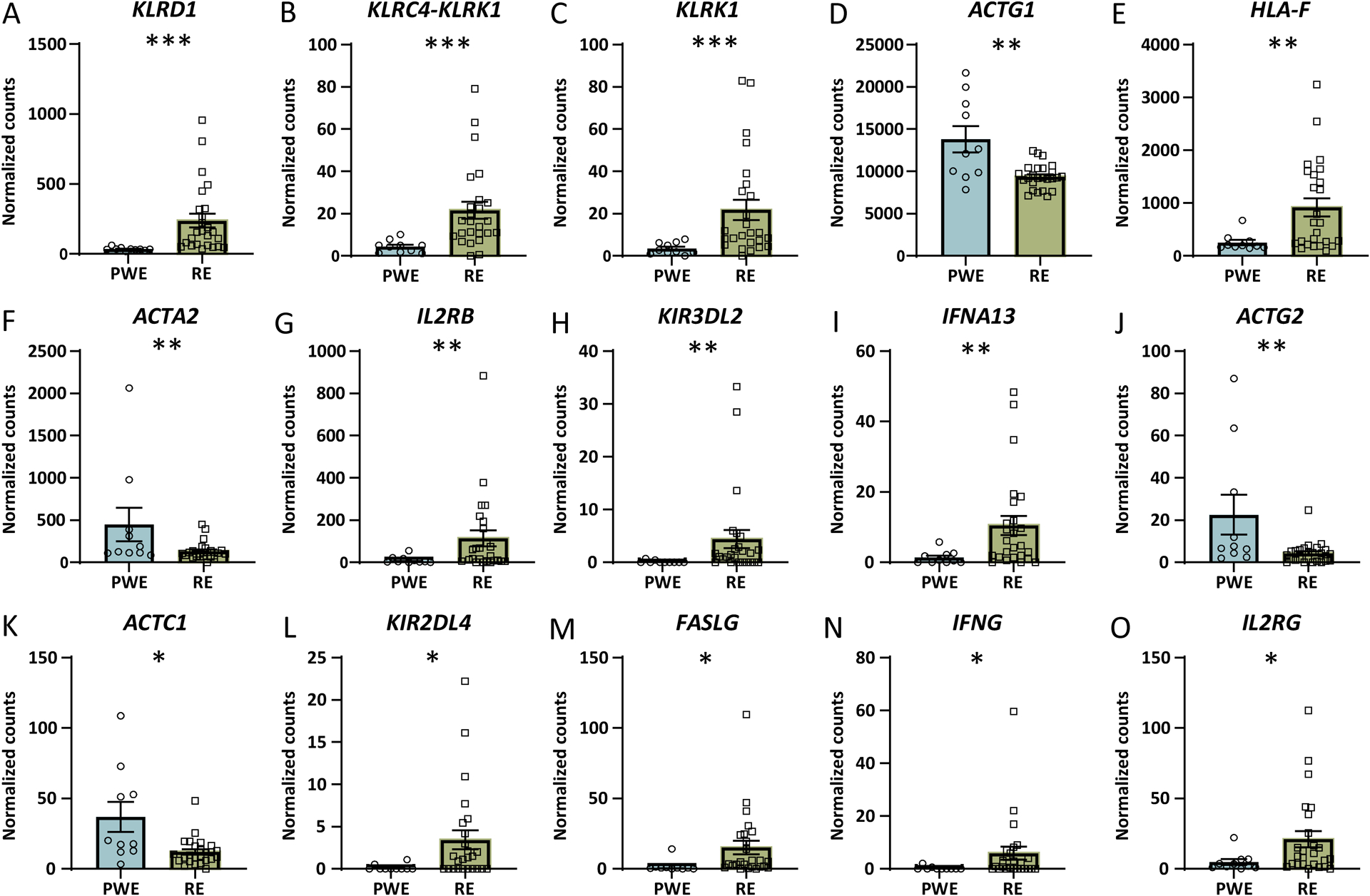
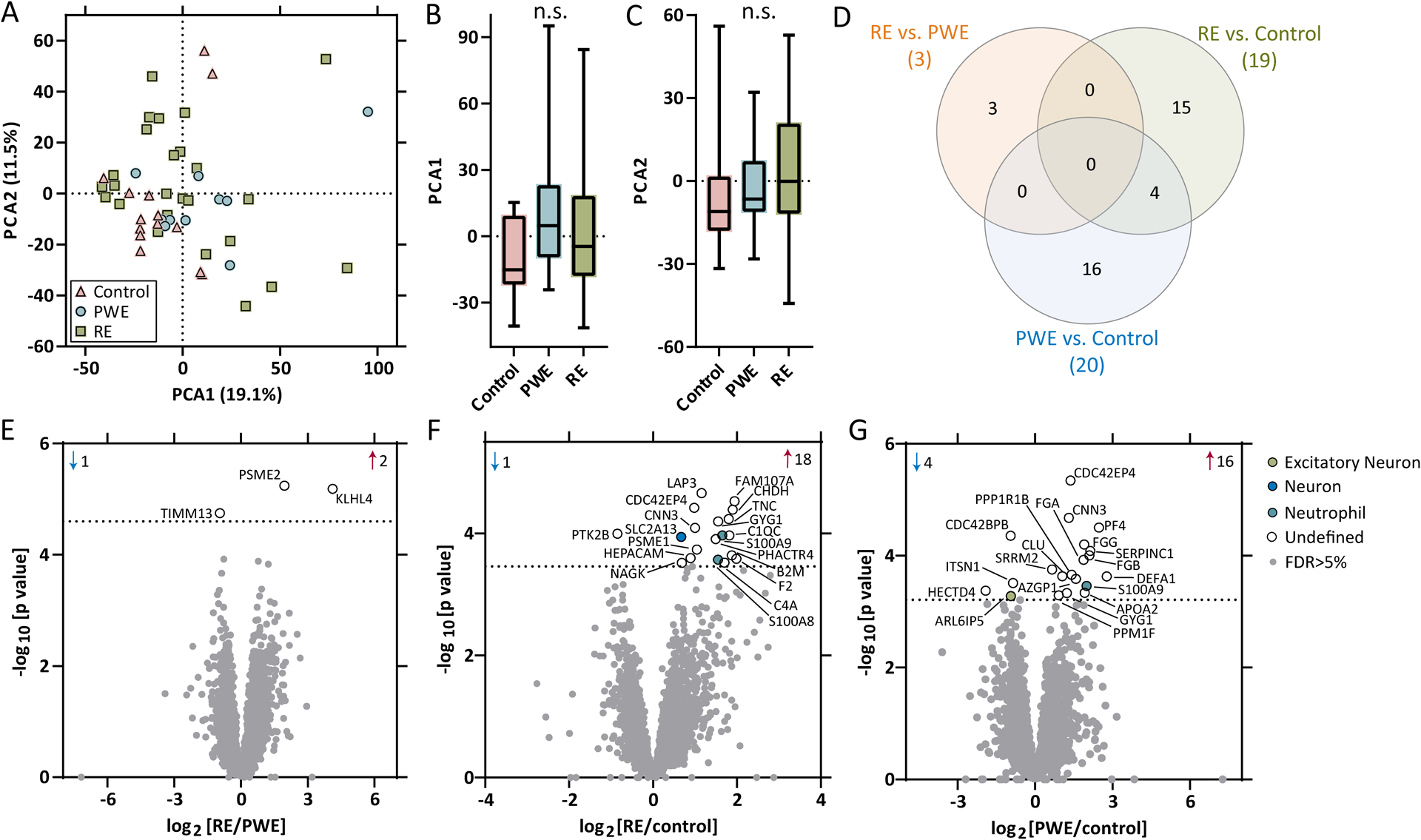
Results: WES revealed no common pathogenic variants in RE, but several rare and likely deleterious variants of unknown significance (VUS; ANGPTL7/MTOR, SCN1A, FCGR3B, MTOR) and more common HLA VUS in >25% of RE cases (HLA-DRB1, HLA-DQA2), all with allele frequency < 5% in the general population. RNAseq in RE versus PWE (1516 altered transcripts) revealed significant activation of crosstalk between dendritic and natural killer cells (p = 7.94 × 10-6 , z = 2.65), in RE versus control (7466 transcripts) neuroinflammation signaling activation (p = 6.31 × 10-13 , z = 5.07), and in PWE versus control (945 transcripts) phagosome formation activation (p = 2.00 × 10-13 , z = 5.61). Proteomics detected fewer altered targets.
Significance: In RE, we identified activated immune signaling pathways and immune cell type annotation enrichment that suggest roles of the innate and adaptive immune responses, as well as HLA variants that may increase vulnerability to RE. Follow-up studies could evaluate cell type density and subregional localization associated with top targets, clinical history (neuropathology, disease duration), and whether modulating crosstalk between dendritic and natural killer cells may limit disease progression.
Keywords: RNAseq; Rasmussen encephalitis; exome; proteomics.
© 2022 International League Against Epilepsy.
Figures
References
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
