

Drosophila melanogaster as a model to study innate immune memory
- PMID: 36338030
- PMCID: PMC9630750
- DOI: 10.3389/fmicb.2022.991678
Drosophila melanogaster as a model to study innate immune memory
Abstract
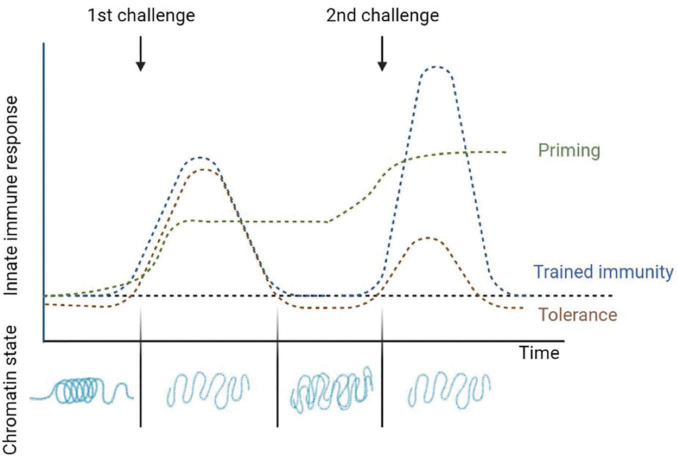
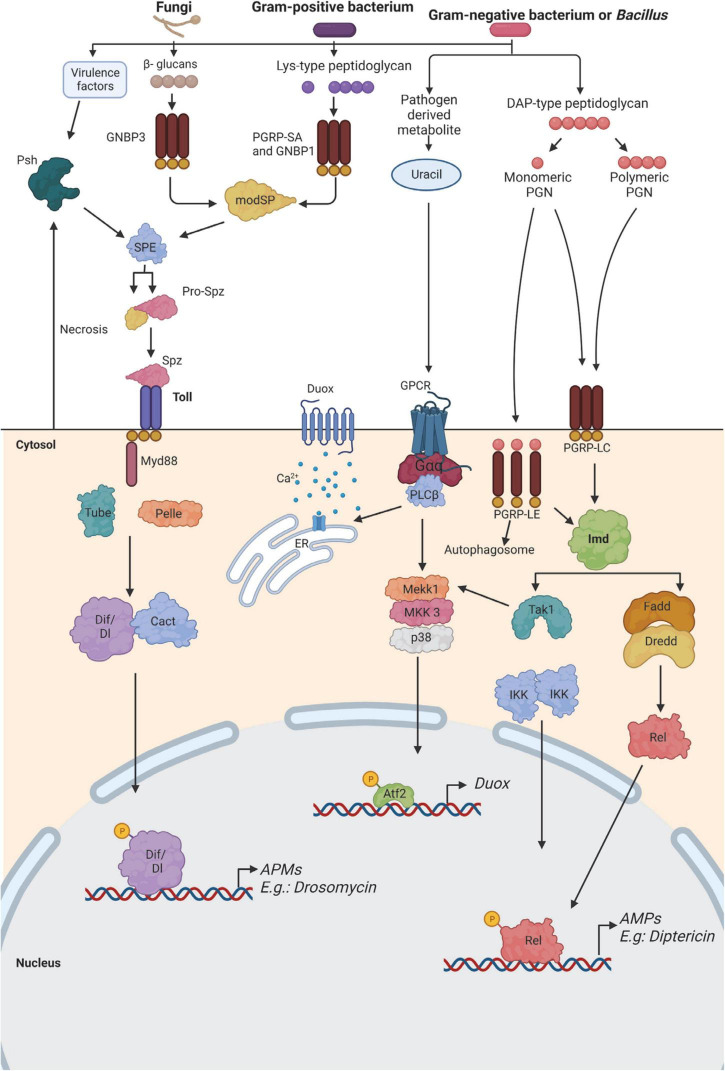
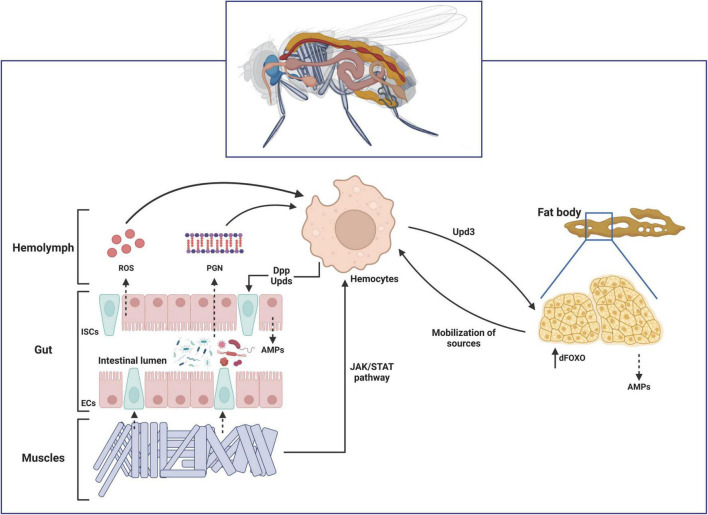
Over the last decades, research regarding innate immune responses has gained increasing importance. A growing body of evidence supports the notion that the innate arm of the immune system could show memory traits. Such traits are thought to be conserved throughout evolution and provide a survival advantage. Several models are available to study these mechanisms. Among them, we find the fruit fly, Drosophila melanogaster. This non-mammalian model has been widely used for innate immune research since it naturally lacks an adaptive response. Here, we aim to review the latest advances in the study of the memory mechanisms of the innate immune response using this animal model.
Keywords: Drosophila melanogaster; infection; innate immune memory; resistance; tolerance; trained immunity.
Copyright © 2022 Arch, Vidal, Koiffman, Melkie and Cardona.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
References
-
- Aaby P., Roth A., Ravn H., Napirna B. M., Rodrigues A., Lisse I. M., et al. (2011). Randomized trial of BCG vaccination at birth to low-birth-weight children: Beneficial nonspecific effects in the neonatal period?. J. Infect. Dis. 204 245–252. - PubMed
-
- Apidianakis Y., Rahme L. G. (2009). Drosophila melanogaster as a model host for studying Pseudomonas aeruginosa infection. Nat. Protoc. 4 1285–1294. - PubMed
Publication types
LinkOut - more resources
Full Text Sources
Molecular Biology Databases