

Mitochondrial dynamics involves molecular and mechanical events in motility, fusion and fission
- PMID: 36340034
- PMCID: PMC9626967
- DOI: 10.3389/fcell.2022.1010232
Mitochondrial dynamics involves molecular and mechanical events in motility, fusion and fission
Abstract
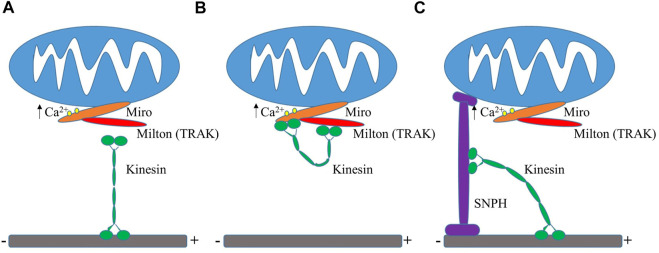
Mitochondria are cell organelles that play pivotal roles in maintaining cell survival, cellular metabolic homeostasis, and cell death. Mitochondria are highly dynamic entities which undergo fusion and fission, and have been shown to be very motile in vivo in neurons and in vitro in multiple cell lines. Fusion and fission are essential for maintaining mitochondrial homeostasis through control of morphology, content exchange, inheritance of mitochondria, maintenance of mitochondrial DNA, and removal of damaged mitochondria by autophagy. Mitochondrial motility occurs through mechanical and molecular mechanisms which translocate mitochondria to sites of high energy demand. Motility also plays an important role in intracellular signaling. Here, we review key features that mediate mitochondrial dynamics and explore methods to advance the study of mitochondrial motility as well as mitochondrial dynamics-related diseases and mitochondrial-targeted therapeutics.
Keywords: disease; fission; fusion; live-cell imaging; mitochondria; mitochondrial DNA; motility; therapeutics.
Copyright © 2022 Green, Hossain and Eckmann.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures


References
-
- Alexander C., Votruba M., Pesch U. E., Thiselton D. L., Mayer S., Moore A., et al. (2000). OPA1, encoding a dynamin-related GTPase, is mutated in autosomal dominant optic atrophy linked to chromosome 3q28. Nat. Genet. 26, 211–215. - PubMed
Publication types
LinkOut - more resources
Full Text Sources