

A plastid nucleoside kinase is involved in inosine salvage and control of purine nucleotide biosynthesis
- PMID: 36342213
- PMCID: PMC9806653
- DOI: 10.1093/plcell/koac320
A plastid nucleoside kinase is involved in inosine salvage and control of purine nucleotide biosynthesis
Abstract
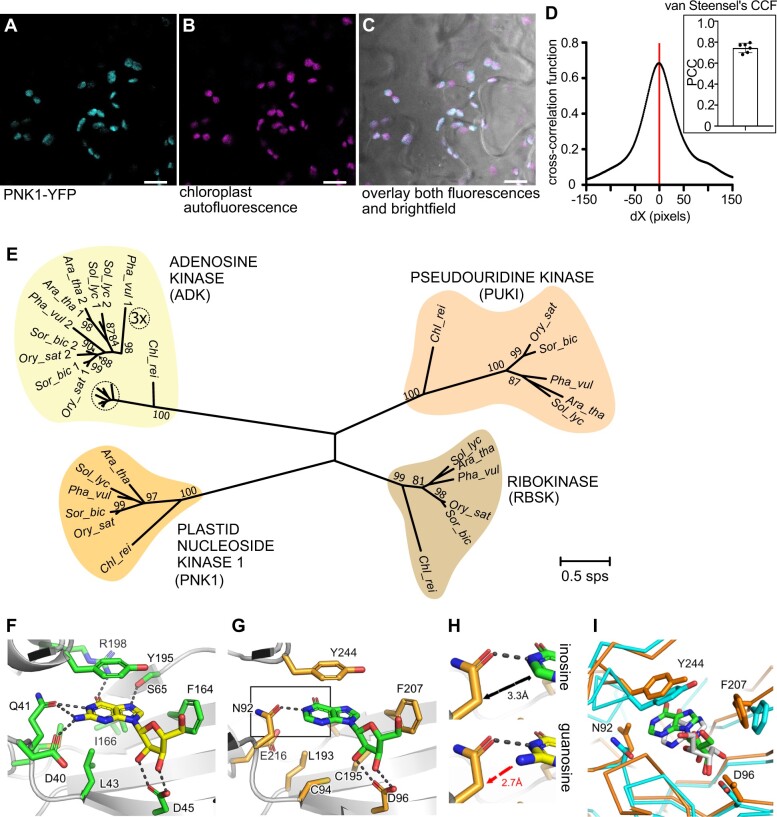
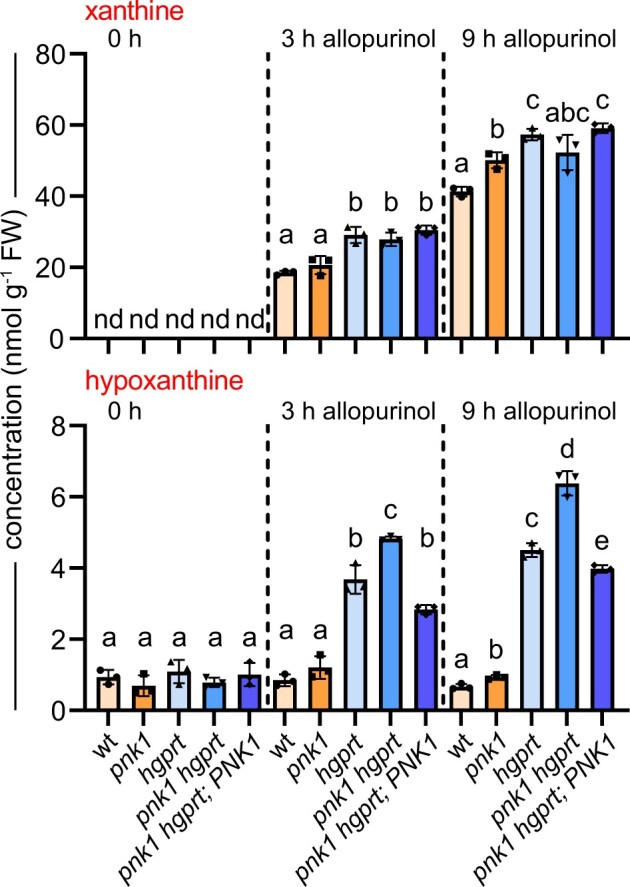
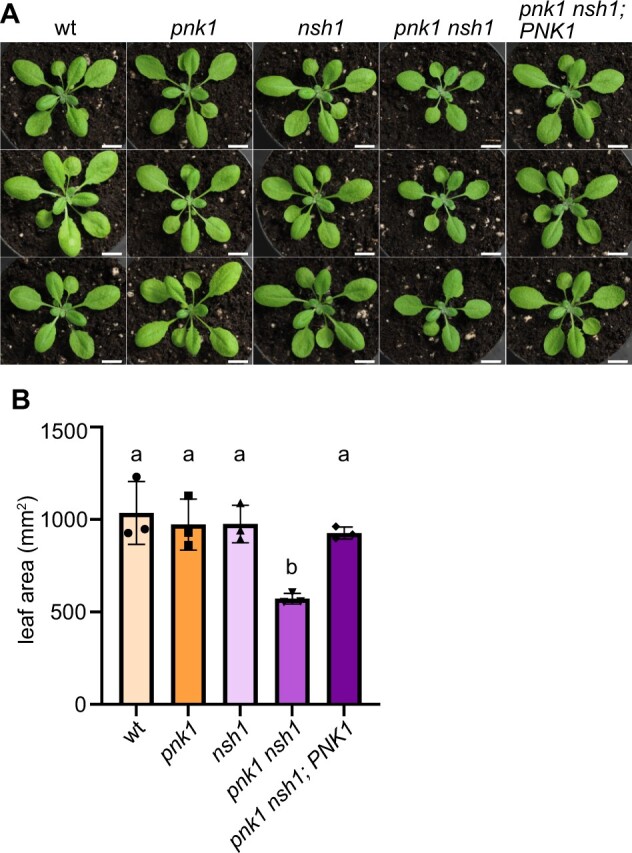
In nucleotide metabolism, nucleoside kinases recycle nucleosides into nucleotides-a process called nucleoside salvage. Nucleoside kinases for adenosine, uridine, and cytidine have been characterized from many organisms, but kinases for inosine and guanosine salvage are not yet known in eukaryotes and only a few such enzymes have been described from bacteria. Here we identified Arabidopsis thaliana PLASTID NUCLEOSIDE KINASE 1 (PNK1), an enzyme highly conserved in plants and green algae belonging to the Phosphofructokinase B family. We demonstrate that PNK1 from A. thaliana is located in plastids and catalyzes the phosphorylation of inosine, 5-aminoimidazole-4-carboxamide-1-β-d-ribose (AICA ribonucleoside), and uridine but not guanosine in vitro, and is involved in inosine salvage in vivo. PNK1 mutation leads to increased flux into purine nucleotide catabolism and, especially in the context of defective uridine degradation, to over-accumulation of uridine and UTP as well as growth depression. The data suggest that PNK1 is involved in feedback regulation of purine nucleotide biosynthesis and possibly also pyrimidine nucleotide biosynthesis. We additionally report that cold stress leads to accumulation of purine nucleotides, probably by inducing nucleotide biosynthesis, but that this adjustment of nucleotide homeostasis to environmental conditions is not controlled by PNK1.
© American Society of Plant Biologists 2022. All rights reserved. For permissions, please email: journals.permissions@oup.com.
Figures






Similar articles
-
Profiles of purine biosynthesis, salvage and degradation in disks of potato (Solanum tuberosum L.) tubers.Planta. 2006 Dec;225(1):115-26. doi: 10.1007/s00425-006-0334-9. Epub 2006 Jul 15. Planta. 2006. PMID: 16845529
-
Two nucleoside uptake systems in Lactococcus lactis: competition between purine nucleosides and cytidine allows for modulation of intracellular nucleotide pools.J Bacteriol. 2003 Mar;185(5):1503-8. doi: 10.1128/JB.185.5.1503-1508.2003. J Bacteriol. 2003. PMID: 12591866 Free PMC article.
-
Key role of uridine kinase and uridine phosphorylase in the homeostatic regulation of purine and pyrimidine salvage in brain.Neurochem Int. 2007 Dec;51(8):517-23. doi: 10.1016/j.neuint.2007.06.007. Epub 2007 Jun 22. Neurochem Int. 2007. PMID: 17643556
-
Purine salvage in plants.Phytochemistry. 2018 Mar;147:89-124. doi: 10.1016/j.phytochem.2017.12.008. Epub 2018 Jan 4. Phytochemistry. 2018. PMID: 29306799 Review.
-
Purine and pyrimidine nucleotide metabolism in higher plants.J Plant Physiol. 2003 Nov;160(11):1271-95. doi: 10.1078/0176-1617-01169. J Plant Physiol. 2003. PMID: 14658380 Review.
Cited by
-
An epiallele of a gene encoding a PfkB-type carbohydrate kinase affects plant architecture in maize.Plant Cell. 2024 Dec 23;37(1):koaf017. doi: 10.1093/plcell/koaf017. Plant Cell. 2024. PMID: 39823309
-
Unveiling the Impact of Organic Fertilizer on Rice (Oryza sativa L.) Salinity Tolerance: Insights from the Integration of NDVI and Metabolomics.Plants (Basel). 2025 Mar 13;14(6):902. doi: 10.3390/plants14060902. Plants (Basel). 2025. PMID: 40265824 Free PMC article.
-
Three Arabidopsis UMP kinases have different roles in pyrimidine nucleotide biosynthesis and (deoxy)CMP salvage.Plant Cell. 2024 Sep 3;36(9):3611-3630. doi: 10.1093/plcell/koae170. Plant Cell. 2024. PMID: 38865437 Free PMC article.
-
Pyrimidine catabolism is required to prevent the accumulation of 5-methyluridine in RNA.Nucleic Acids Res. 2023 Aug 11;51(14):7451-7464. doi: 10.1093/nar/gkad529. Nucleic Acids Res. 2023. PMID: 37334828 Free PMC article.
-
Nucleotide Imbalance, Provoked by Downregulation of Aspartate Transcarbamoylase Impairs Cold Acclimation in Arabidopsis.Molecules. 2023 Feb 7;28(4):1585. doi: 10.3390/molecules28041585. Molecules. 2023. PMID: 36838573 Free PMC article.
References
-
- Allen M, Qin WS, Moreau F, Moffatt B (2002) Adenine phosphoribosyltransferase isoforms of Arabidopsis and their potential contributions to adenine and cytokinin metabolism. Physiol Plantarum 115: 56–68 - PubMed
-
- Alonso JM, Stepanova AN, Leisse TJ, Kim CJ, Chen HM, Shinn P, Stevenson DK, Zimmerman J, Barajas P, Cheuk R, et al. (2003) Genome-wide insertional mutagenesis of Arabidopsis thaliana. Science 301: 653–657 - PubMed
-
- Ashihara H, Stasolla C, Fujimura T, Crozier A (2018) Purine salvage in plants. Phytochem 147: 89–124 - PubMed
-
- Baena-Gonzalez E, Rolland F, Thevelein JM, Sheen J (2007) A central integrator of transcription networks in plant stress and energy signalling. Nature 448: 938–942 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
