

Origins and Evolution of Human Tandem Duplicated Exon Substitution Events
- PMID: 36346145
- PMCID: PMC9741552
- DOI: 10.1093/gbe/evac162
Origins and Evolution of Human Tandem Duplicated Exon Substitution Events
Abstract
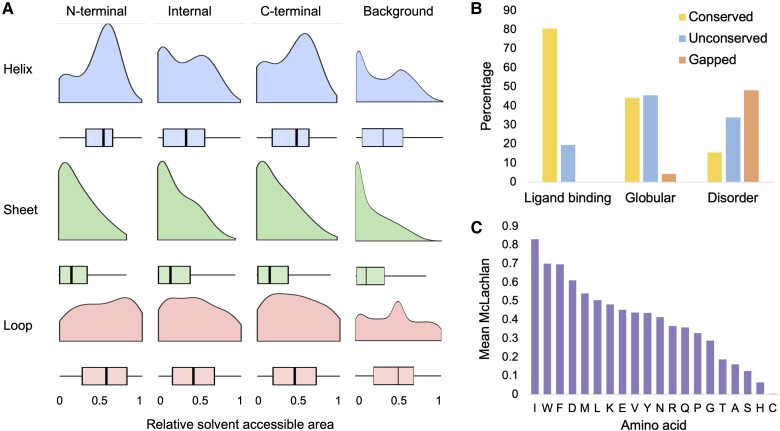
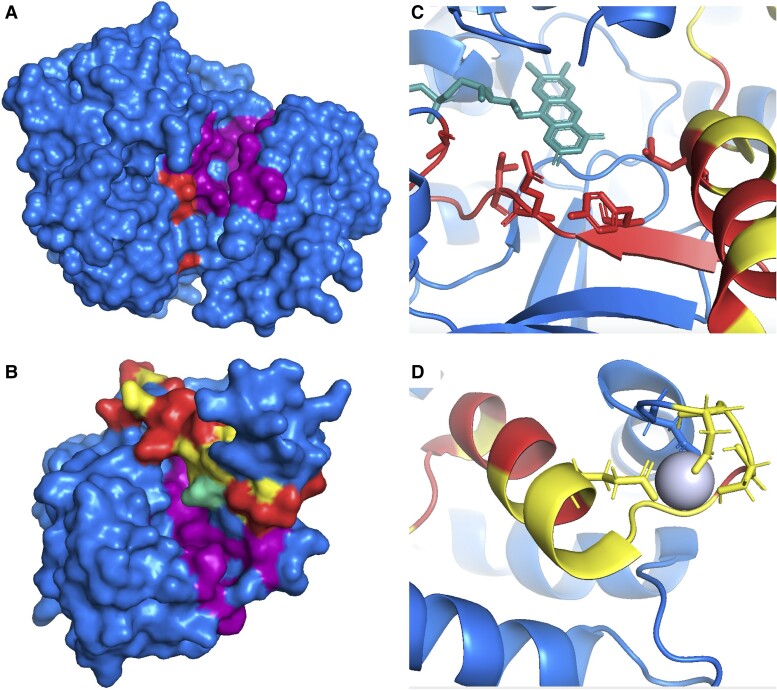
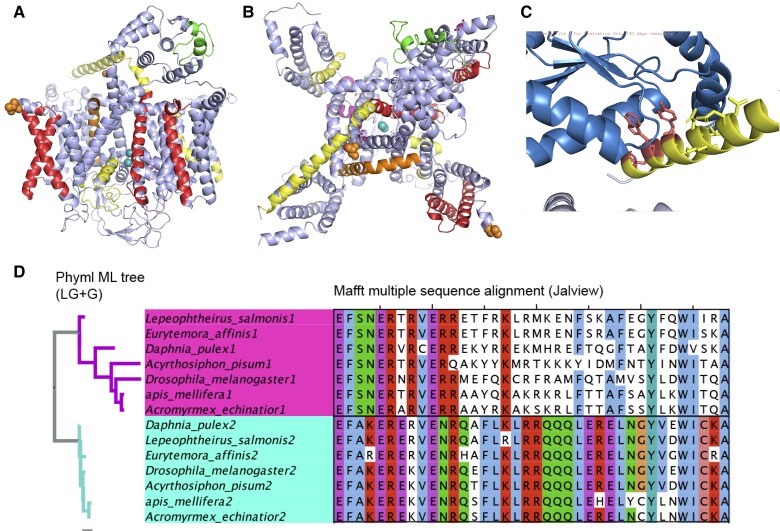
The mutually exclusive splicing of tandem duplicated exons produces protein isoforms that are identical save for a homologous region that allows for the fine tuning of protein function. Tandem duplicated exon substitution events are rare, yet highly important alternative splicing events. Most events are ancient, their isoforms are highly expressed, and they have significantly more pathogenic mutations than other splice events. Here, we analyzed the physicochemical properties and functional roles of the homologous polypeptide regions produced by the 236 tandem duplicated exon substitutions annotated in the human gene set. We find that the most important structural and functional residues in these homologous regions are maintained, and that most changes are conservative rather than drastic. Three quarters of the isoforms produced from tandem duplicated exon substitution events are tissue-specific, particularly in nervous and cardiac tissues, and tandem duplicated exon substitution events are enriched in functional terms related to structures in the brain and skeletal muscle. We find considerable evidence for the convergent evolution of tandem duplicated exon substitution events in vertebrates, arthropods, and nematodes. Twelve human gene families have orthologues with tandem duplicated exon substitution events in both Drosophila melanogaster and Caenorhabditis elegans. Six of these gene families are ion transporters, suggesting that tandem exon duplication in genes that control the flow of ions into the cell has an adaptive benefit. The ancient origins, the strong indications of tissue-specific functions, and the evidence of convergent evolution suggest that these events may have played important roles in the evolution of animal tissues and organs.
Keywords: alternative splicing; exon duplication; function; ion channels; proteomics; tissue specificity.
© The Author(s) 2022. Published by Oxford University Press on behalf of Society for Molecular Biology and Evolution.
Figures


Similar articles
-
The clinical importance of tandem exon duplication-derived substitutions.Nucleic Acids Res. 2021 Aug 20;49(14):8232-8246. doi: 10.1093/nar/gkab623. Nucleic Acids Res. 2021. PMID: 34302486 Free PMC article.
-
Alternatively Spliced Homologous Exons Have Ancient Origins and Are Highly Expressed at the Protein Level.PLoS Comput Biol. 2015 Jun 10;11(6):e1004325. doi: 10.1371/journal.pcbi.1004325. eCollection 2015 Jun. PLoS Comput Biol. 2015. PMID: 26061177 Free PMC article.
-
Common exon duplication in animals and its role in alternative splicing.Hum Mol Genet. 2002 Jun 15;11(13):1561-7. doi: 10.1093/hmg/11.13.1561. Hum Mol Genet. 2002. PMID: 12045209
-
Evolutionary convergence of alternative splicing in ion channels.Trends Genet. 2004 Apr;20(4):171-6. doi: 10.1016/j.tig.2004.02.001. Trends Genet. 2004. PMID: 15101391 Review.
-
Functional regulation of alternatively spliced Na+/Ca2+ exchanger (NCX1) isoforms.Ann N Y Acad Sci. 2002 Nov;976:187-96. doi: 10.1111/j.1749-6632.2002.tb04740.x. Ann N Y Acad Sci. 2002. PMID: 12502560 Review.
Cited by
-
Exonize: a tool for finding and classifying exon duplications in annotated genomes.Bioinform Adv. 2025 Jul 28;5(1):vbaf177. doi: 10.1093/bioadv/vbaf177. eCollection 2025. Bioinform Adv. 2025. PMID: 40799492 Free PMC article.
-
Evidence for widespread translation of 5' untranslated regions.Nucleic Acids Res. 2024 Aug 12;52(14):8112-8126. doi: 10.1093/nar/gkae571. Nucleic Acids Res. 2024. PMID: 38953162 Free PMC article.
-
Toward a comprehensive profiling of alternative splicing proteoform structures, interactions and functions.Curr Opin Struct Biol. 2025 Feb;90:102979. doi: 10.1016/j.sbi.2024.102979. Epub 2025 Jan 7. Curr Opin Struct Biol. 2025. PMID: 39778413 Free PMC article. Review.
-
HMZDupFinder: a robust computational approach for detecting intragenic homozygous duplications from exome sequencing data.Nucleic Acids Res. 2024 Feb 28;52(4):e18. doi: 10.1093/nar/gkad1223. Nucleic Acids Res. 2024. PMID: 38153174 Free PMC article.
-
A deep audit of the PeptideAtlas database uncovers evidence for unannotated coding genes and aberrant translation.bioRxiv [Preprint]. 2024 Nov 15:2024.11.14.623419. doi: 10.1101/2024.11.14.623419. bioRxiv. 2024. PMID: 39605392 Free PMC article. Preprint.
References
-
- Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ. 1990. Basic local alignment search tool. J Mol Biol. 215:403–410. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
