

Deficiency for SAMHD1 activates MDA5 in a cGAS/STING-dependent manner
- PMID: 36346347
- PMCID: PMC9648672
- DOI: 10.1084/jem.20220829
Deficiency for SAMHD1 activates MDA5 in a cGAS/STING-dependent manner
Abstract
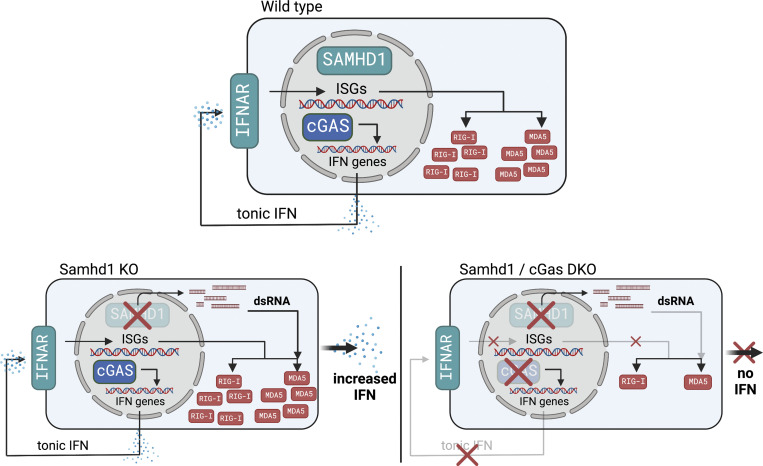
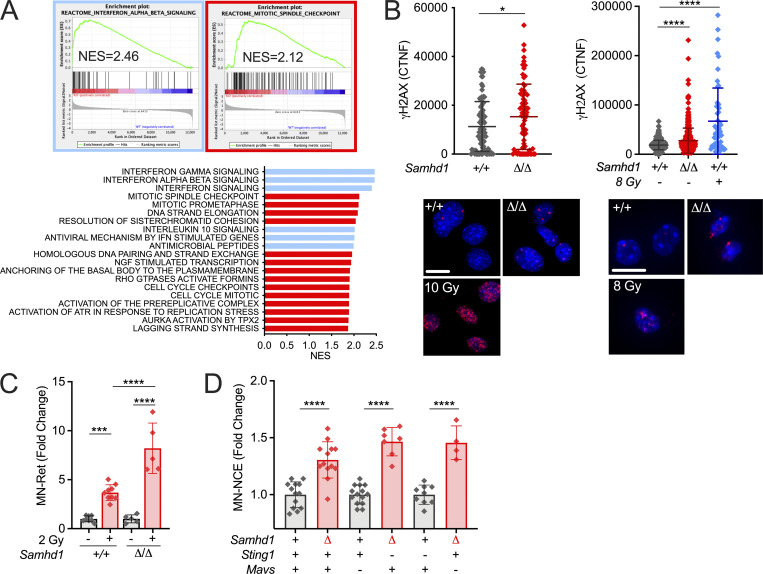
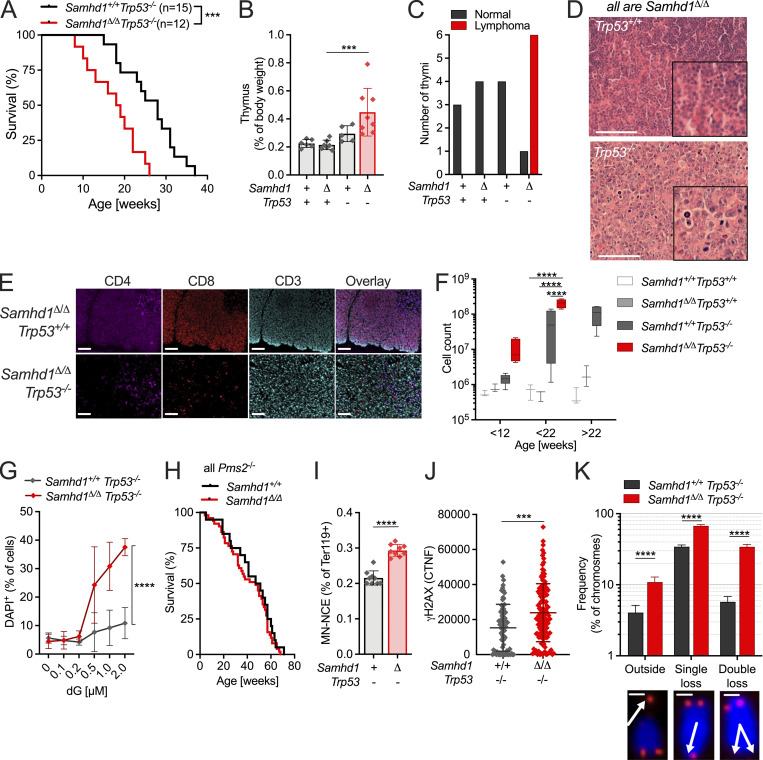
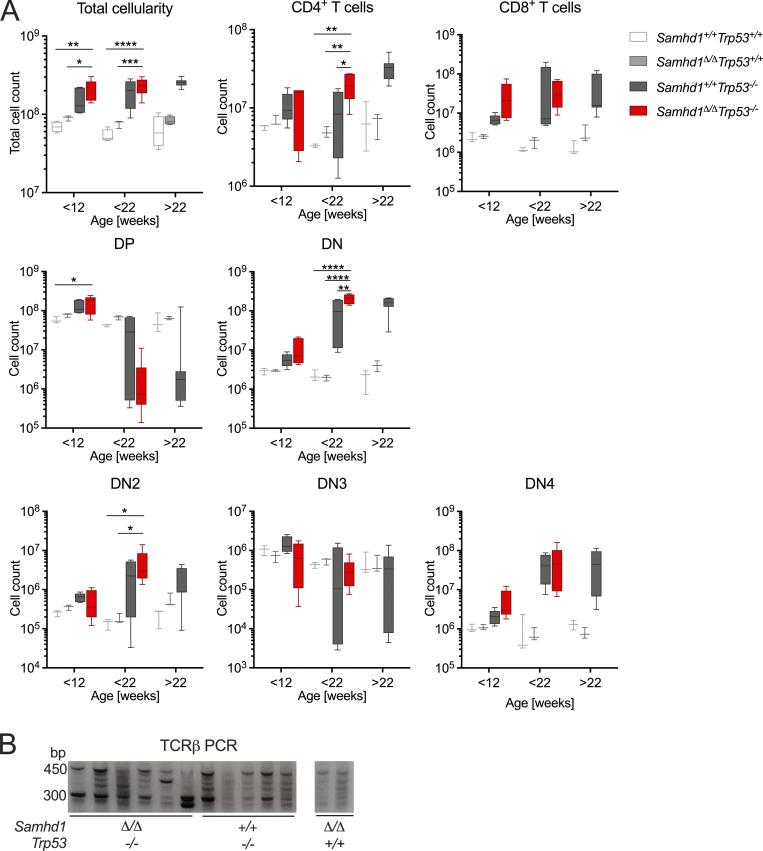
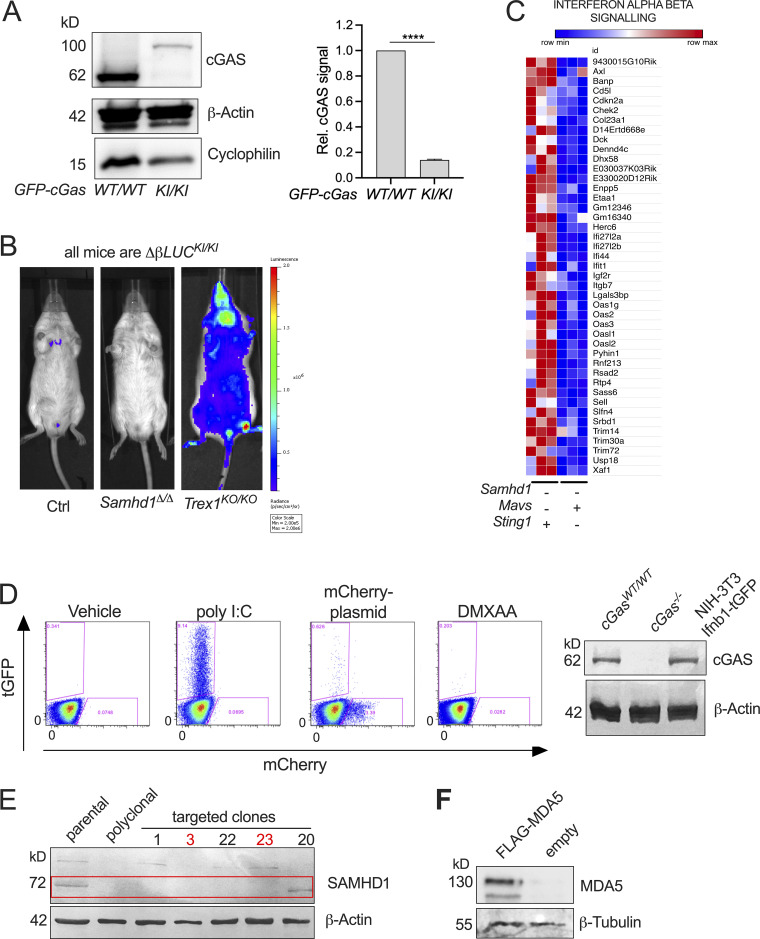
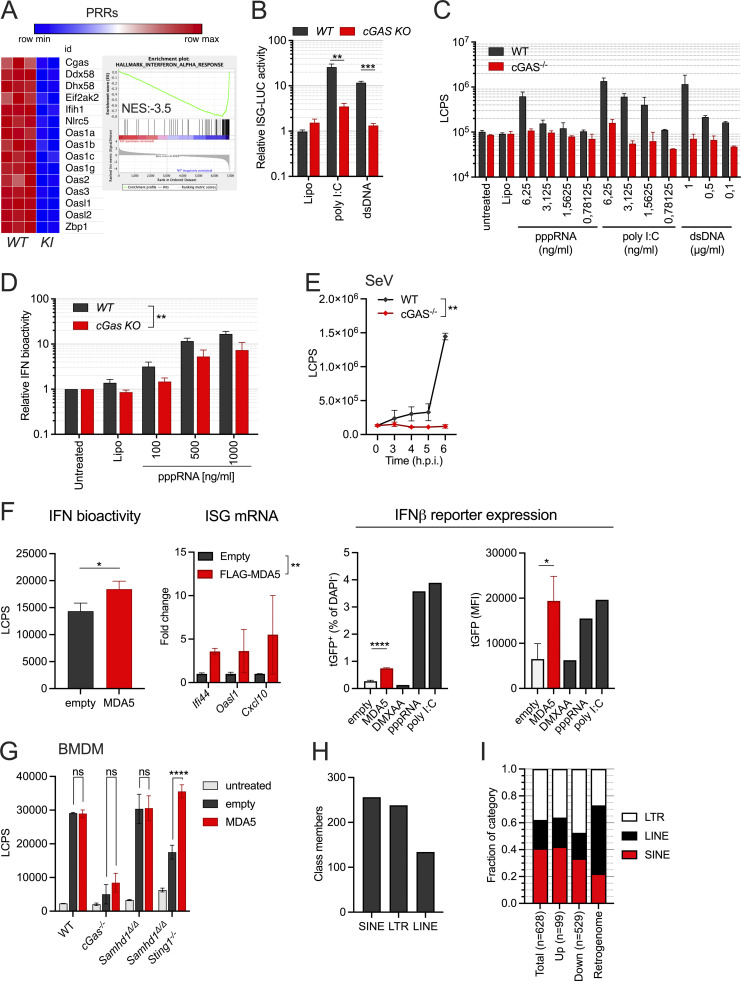
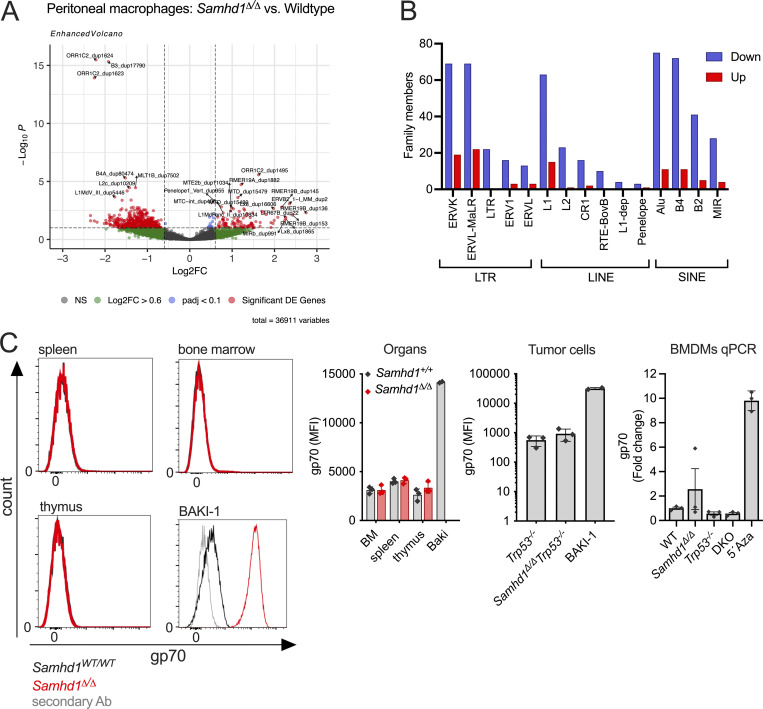
Defects in nucleic acid metabolizing enzymes can lead to spontaneous but selective activation of either cGAS/STING or RIG-like receptor (RLR) signaling, causing type I interferon-driven inflammatory diseases. In these pathophysiological conditions, activation of the DNA sensor cGAS and IFN production are linked to spontaneous DNA damage. Physiological, or tonic, IFN signaling on the other hand is essential to functionally prime nucleic acid sensing pathways. Here, we show that low-level chronic DNA damage in mice lacking the Aicardi-Goutières syndrome gene SAMHD1 reduced tumor-free survival when crossed to a p53-deficient, but not to a DNA mismatch repair-deficient background. Increased DNA damage did not result in higher levels of type I interferon. Instead, we found that the chronic interferon response in SAMHD1-deficient mice was driven by the MDA5/MAVS pathway but required functional priming through the cGAS/STING pathway. Our work positions cGAS/STING upstream of tonic IFN signaling in Samhd1-deficient mice and highlights an important role of the pathway in physiological and pathophysiological innate immune priming.
© 2022 Schumann et al.
Conflict of interest statement
Disclosures: R. Behrendt reported grants from Hoffmann-La Roche, ISD Immunotech, and IFM Therapeutics outside the submitted work. No other disclosures were reported.
Figures
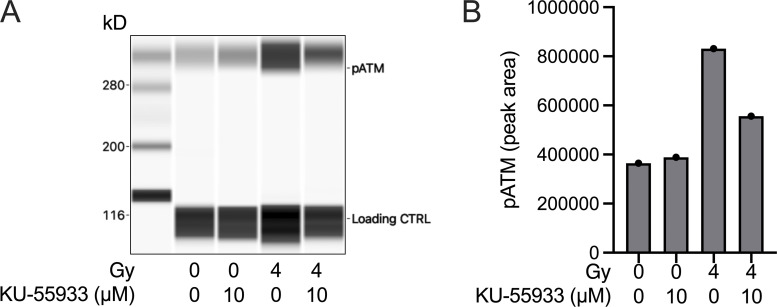
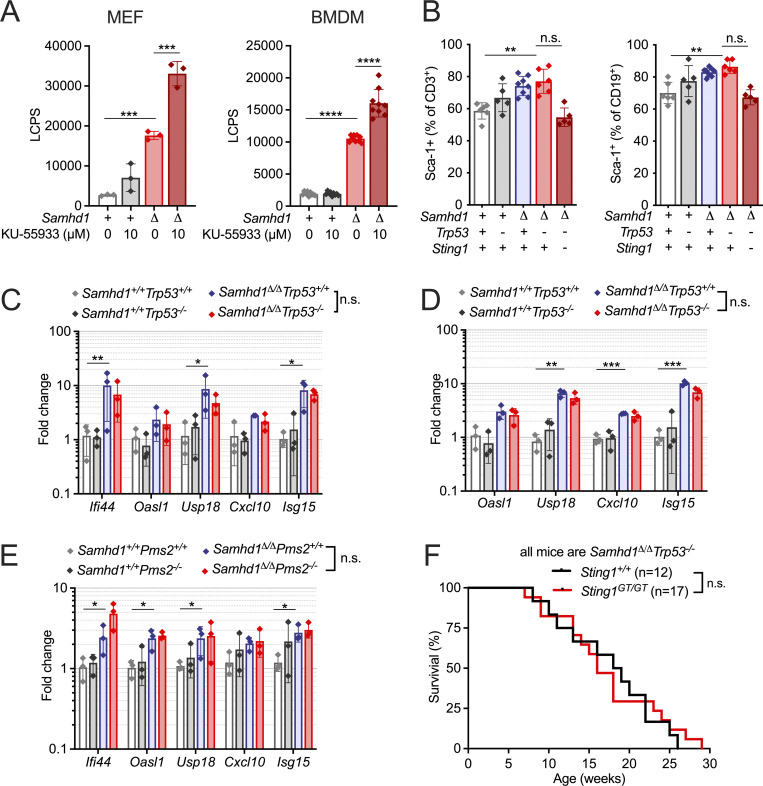
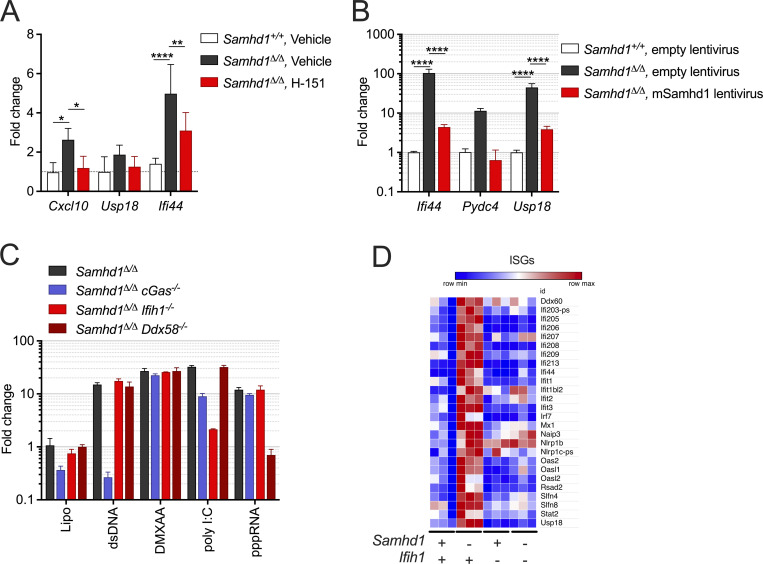
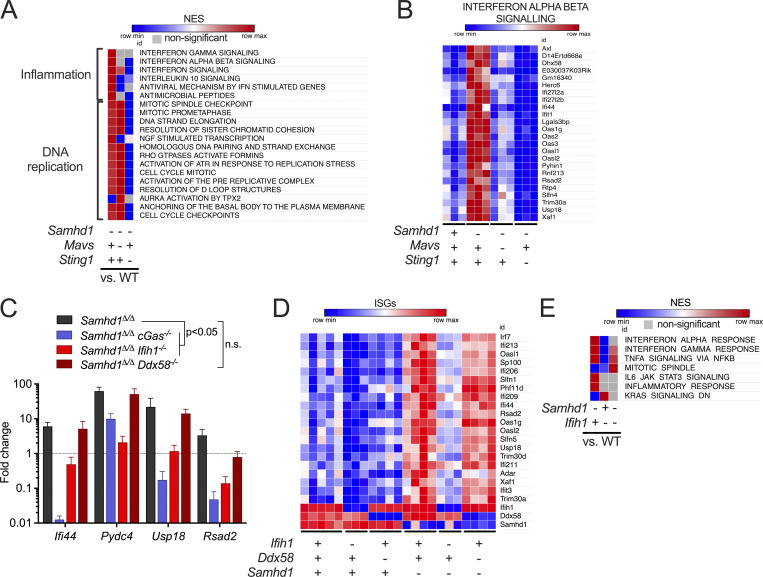
References
-
- Aditi, S.M., Downing S.M., Schreiner P.A., Kwak Y.D., Li Y., Shaw T.I., Russell H.R., and McKinnon P.J.. 2021. Genome instability independent of type I interferon signaling drives neuropathology caused by impaired ribonucleotide excision repair. Neuron. 109:3962–3979.e6. 10.1016/j.neuron.2021.09.040 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
