

wnt16 regulates spine and muscle morphogenesis through parallel signals from notochord and dermomyotome
- PMID: 36346812
- PMCID: PMC9674140
- DOI: 10.1371/journal.pgen.1010496
wnt16 regulates spine and muscle morphogenesis through parallel signals from notochord and dermomyotome
Abstract
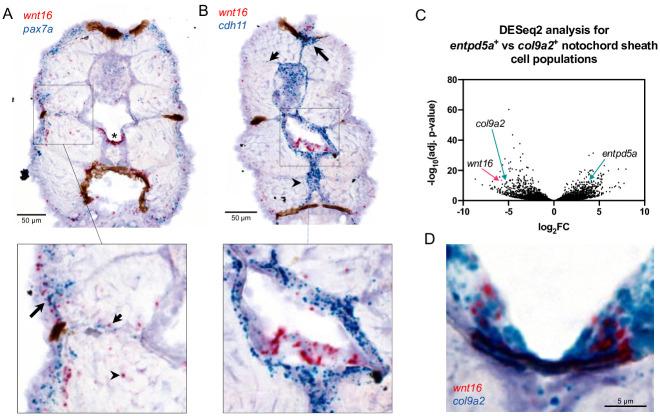
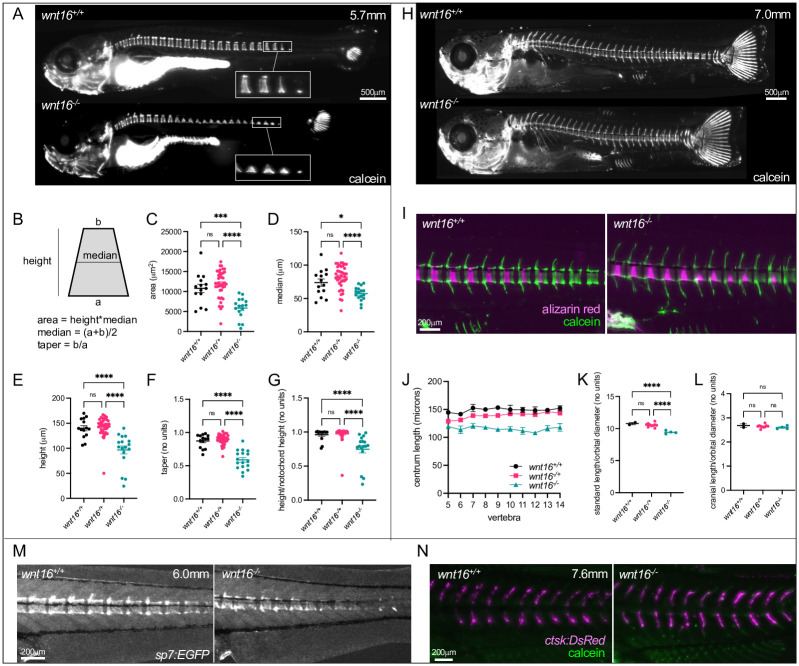
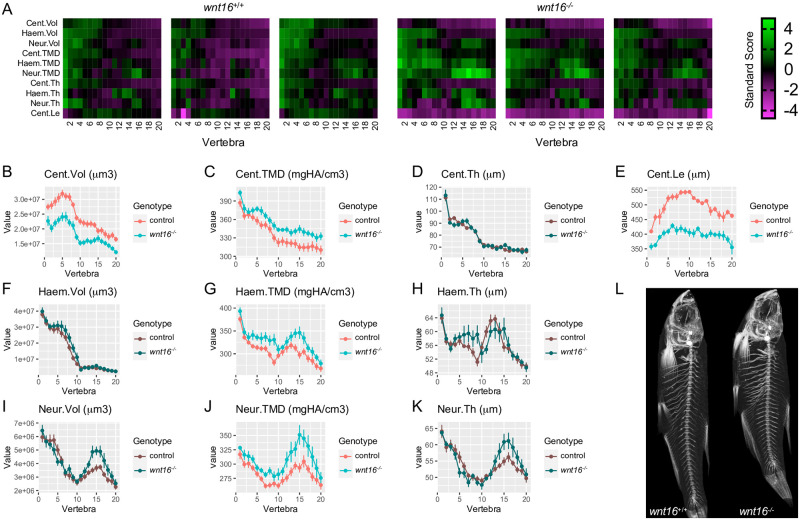
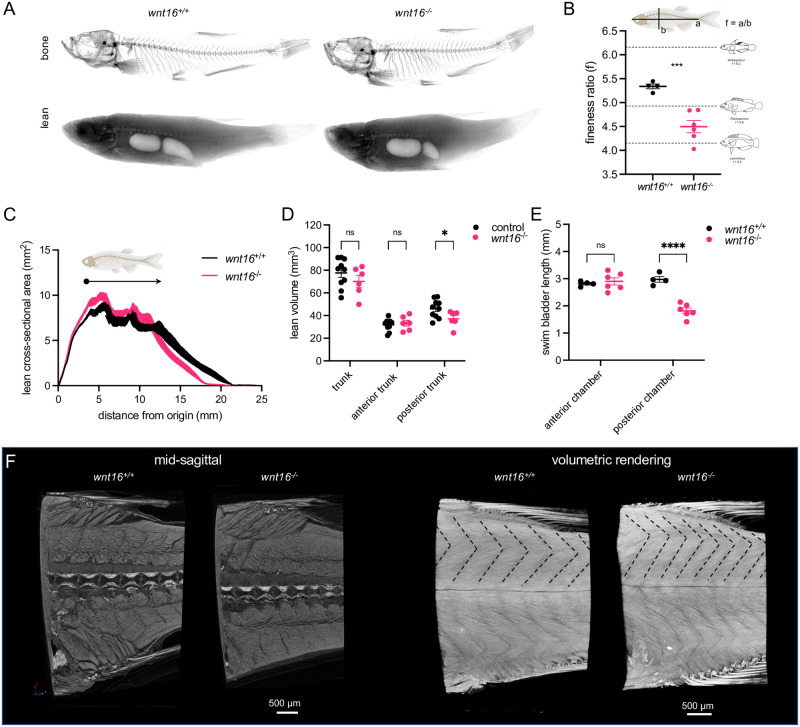
Bone and muscle are coupled through developmental, mechanical, paracrine, and autocrine signals. Genetic variants at the CPED1-WNT16 locus are dually associated with bone- and muscle-related traits. While Wnt16 is necessary for bone mass and strength, this fails to explain pleiotropy at this locus. Here, we show wnt16 is required for spine and muscle morphogenesis in zebrafish. In embryos, wnt16 is expressed in dermomyotome and developing notochord, and contributes to larval myotome morphology and notochord elongation. Later, wnt16 is expressed at the ventral midline of the notochord sheath, and contributes to spine mineralization and osteoblast recruitment. Morphological changes in wnt16 mutant larvae are mirrored in adults, indicating that wnt16 impacts bone and muscle morphology throughout the lifespan. Finally, we show that wnt16 is a gene of major effect on lean mass at the CPED1-WNT16 locus. Our findings indicate that Wnt16 is secreted in structures adjacent to developing bone (notochord) and muscle (dermomyotome) where it affects the morphogenesis of each tissue, thereby rendering wnt16 expression into dual effects on bone and muscle morphology. This work expands our understanding of wnt16 in musculoskeletal development and supports the potential for variants to act through WNT16 to influence bone and muscle via parallel morphogenetic processes.
Copyright: © 2022 Watson et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Multiple Mechanisms Explain Genetic Effects at the CPED1-WNT16 Bone Mineral Density Locus.Curr Osteoporos Rep. 2023 Apr;21(2):173-183. doi: 10.1007/s11914-023-00783-w. Epub 2023 Mar 21. Curr Osteoporos Rep. 2023. PMID: 36943599 Free PMC article. Review.
-
Notochordal Signals Establish Phylogenetic Identity of the Teleost Spine.Curr Biol. 2020 Jul 20;30(14):2805-2814.e3. doi: 10.1016/j.cub.2020.05.037. Epub 2020 Jun 18. Curr Biol. 2020. PMID: 32559448 Free PMC article.
-
Loss of Wnt16 Leads to Skeletal Deformities and Downregulation of Bone Developmental Pathway in Zebrafish.Int J Mol Sci. 2021 Jun 22;22(13):6673. doi: 10.3390/ijms22136673. Int J Mol Sci. 2021. PMID: 34206401 Free PMC article.
-
[Early stages of myogenesis as seen through the action of the myf-5 gene].C R Seances Soc Biol Fil. 1997;191(1):43-54. C R Seances Soc Biol Fil. 1997. PMID: 9181127 French.
-
Wnt16 signaling in bone homeostasis and osteoarthristis.Front Endocrinol (Lausanne). 2022 Dec 22;13:1095711. doi: 10.3389/fendo.2022.1095711. eCollection 2022. Front Endocrinol (Lausanne). 2022. PMID: 36619549 Free PMC article. Review.
Cited by
-
Zebrafish as a Model for Osteoporosis: Functional Validations of Genome-Wide Association Studies.Curr Osteoporos Rep. 2023 Dec;21(6):650-659. doi: 10.1007/s11914-023-00831-5. Epub 2023 Nov 16. Curr Osteoporos Rep. 2023. PMID: 37971665 Review.
-
Loss of cped1 does not affect bone and lean tissue in zebrafish.JBMR Plus. 2024 Dec 10;9(2):ziae159. doi: 10.1093/jbmrpl/ziae159. eCollection 2025 Feb. JBMR Plus. 2024. PMID: 39776615 Free PMC article.
-
The Cys-His-Gly triplet within the WNT motif is essential for Wnt16 function in vivo.MicroPubl Biol. 2025 Aug 8;2025:10.17912/micropub.biology.001736. doi: 10.17912/micropub.biology.001736. eCollection 2025. MicroPubl Biol. 2025. PMID: 40861003 Free PMC article.
-
Loss of cped1 does not affect bone and lean tissue in zebrafish.bioRxiv [Preprint]. 2024 Aug 17:2024.07.10.601974. doi: 10.1101/2024.07.10.601974. bioRxiv. 2024. Update in: JBMR Plus. 2024 Dec 10;9(2):ziae159. doi: 10.1093/jbmrpl/ziae159. PMID: 39026892 Free PMC article. Updated. Preprint.
-
Standardization of bone morphometry and mineral density assessments in zebrafish and other small laboratory fishes using X-ray radiography and micro-computed tomography.J Bone Miner Res. 2024 Nov 29;39(12):1695-1710. doi: 10.1093/jbmr/zjae171. J Bone Miner Res. 2024. PMID: 39475005 Free PMC article.
References
-
- Crepaldi G, Maggi S. Sarcopenia and osteoporosis: A hazardous duet. J Endocrinol Invest. 2005;28(10 Suppl):66–8. Epub 2006/03/23. . - PubMed
-
- Medina-Gomez C, Kemp JP, Dimou NL, Kreiner E, Chesi A, Zemel BS, et al.. Bivariate genome-wide association meta-analysis of pediatric musculoskeletal traits reveals pleiotropic effects at the SREBF1/TOM1L2 locus. Nat Commun. 2017;8(1):121. Epub 2017/07/27. doi: 10.1038/s41467-017-00108-3 . - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
