

Structural basis for sequence-independent substrate selection by eukaryotic wobble base tRNA deaminase ADAT2/3
- PMID: 36347890
- PMCID: PMC9643335
- DOI: 10.1038/s41467-022-34441-z
Structural basis for sequence-independent substrate selection by eukaryotic wobble base tRNA deaminase ADAT2/3
Abstract
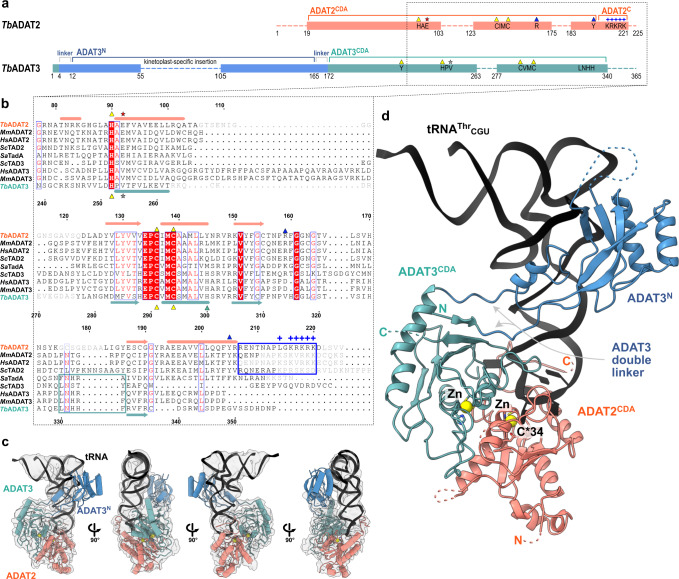
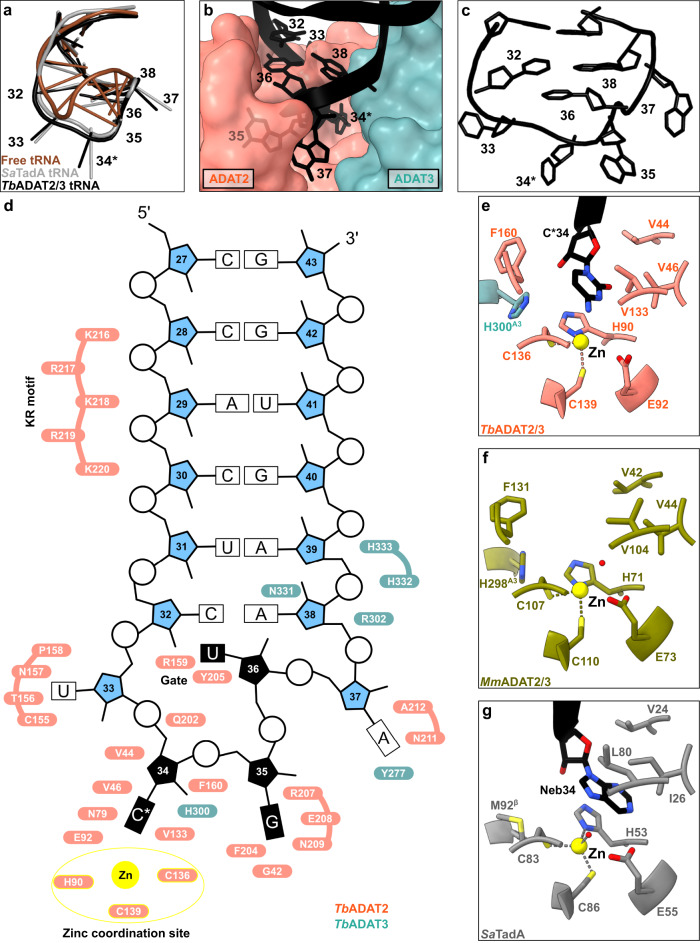
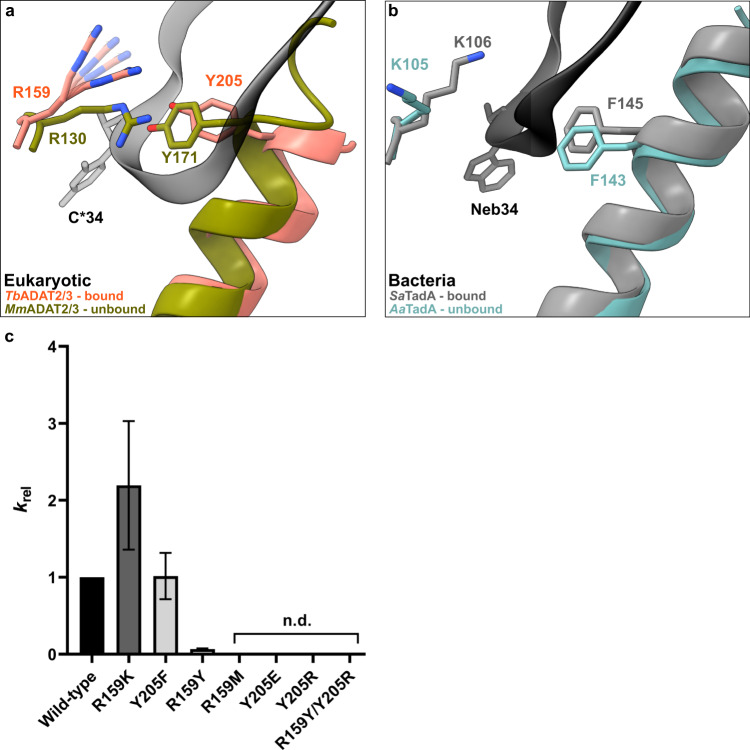
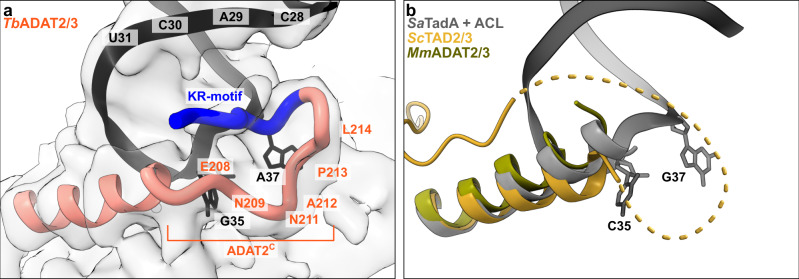
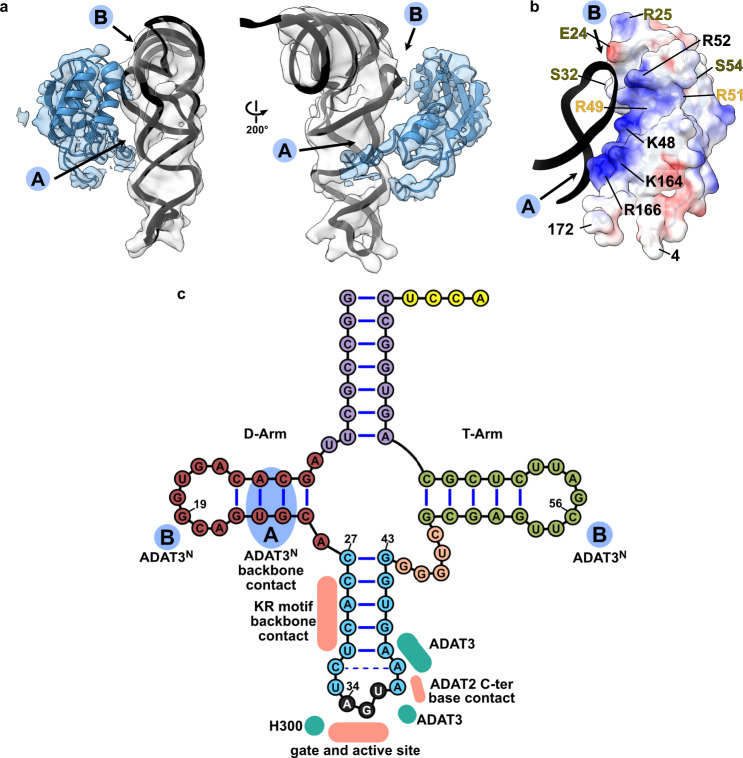
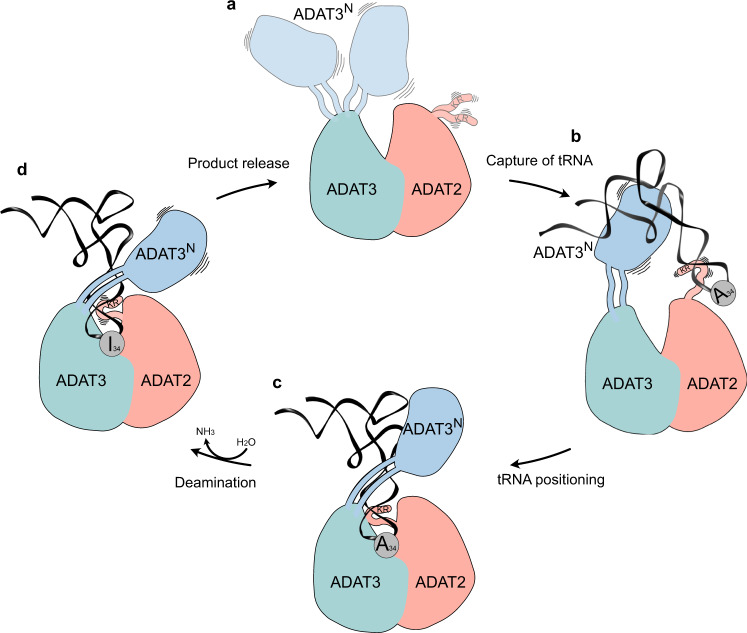
The essential deamination of adenosine A34 to inosine at the wobble base is the individual tRNA modification with the greatest effects on mRNA decoding, empowering a single tRNA to translate three different codons. To date, many aspects of how eukaryotic deaminases specifically select their multiple substrates remain unclear. Here, using cryo-EM, we present the structure of a eukaryotic ADAT2/3 deaminase bound to a full-length tRNA, revealing that the enzyme distorts the anticodon loop, but in contrast to the bacterial enzymes, selects its substrate via sequence-independent contacts of eukaryote-acquired flexible or intrinsically unfolded motifs distal from the conserved catalytic core. A gating mechanism for substrate entry to the active site is identified. Our multi-step tRNA recognition model yields insights into how RNA editing by A34 deamination evolved, shaped the genetic code, and directly impacts the eukaryotic proteome.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
The structure of the mouse ADAT2/ADAT3 complex reveals the molecular basis for mammalian tRNA wobble adenosine-to-inosine deamination.Nucleic Acids Res. 2021 Jun 21;49(11):6529-6548. doi: 10.1093/nar/gkab436. Nucleic Acids Res. 2021. PMID: 34057470 Free PMC article.
-
Biochemical and structural studies of A-to-I editing by tRNA:A34 deaminases at the wobble position of transfer RNA.Biochemistry. 2005 Sep 13;44(36):12057-65. doi: 10.1021/bi050499f. Biochemistry. 2005. PMID: 16142903
-
Identification and rescue of a tRNA wobble inosine deficiency causing intellectual disability disorder.RNA. 2020 Nov;26(11):1654-1666. doi: 10.1261/rna.076380.120. Epub 2020 Aug 6. RNA. 2020. PMID: 32763916 Free PMC article.
-
Factors That Shape Eukaryotic tRNAomes: Processing, Modification and Anticodon-Codon Use.Biomolecules. 2017 Mar 8;7(1):26. doi: 10.3390/biom7010026. Biomolecules. 2017. PMID: 28282871 Free PMC article. Review.
-
Enzymatic conversion of adenosine to inosine and to N1-methylinosine in transfer RNAs: a review.Biochimie. 1996;78(6):488-501. doi: 10.1016/0300-9084(96)84755-9. Biochimie. 1996. PMID: 8915538 Review.
Cited by
-
Unveiling the A-to-I mRNA editing machinery and its regulation and evolution in fungi.Nat Commun. 2024 May 10;15(1):3934. doi: 10.1038/s41467-024-48336-8. Nat Commun. 2024. PMID: 38729938 Free PMC article.
-
Structural basis of tRNA recognition by the m3C RNA methyltransferase METTL6 in complex with SerRS seryl-tRNA synthetase.Nat Struct Mol Biol. 2024 Oct;31(10):1614-1624. doi: 10.1038/s41594-024-01341-3. Epub 2024 Jun 25. Nat Struct Mol Biol. 2024. PMID: 38918637 Free PMC article.
-
Inosine induces stemness features in CAR-T cells and enhances potency.Cancer Cell. 2024 Feb 12;42(2):266-282.e8. doi: 10.1016/j.ccell.2024.01.002. Epub 2024 Jan 25. Cancer Cell. 2024. PMID: 38278150 Free PMC article.
-
RNA editing enzymes: structure, biological functions and applications.Cell Biosci. 2024 Mar 16;14(1):34. doi: 10.1186/s13578-024-01216-6. Cell Biosci. 2024. PMID: 38493171 Free PMC article. Review.
-
A large-scale curated and filterable dataset for cryo-EM foundation model pre-training.Sci Data. 2025 Jun 7;12(1):960. doi: 10.1038/s41597-025-05179-2. Sci Data. 2025. PMID: 40483273 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
