

BLIMP1 and NR4A3 transcription factors reciprocally regulate antitumor CAR T cell stemness and exhaustion
- PMID: 36350986
- PMCID: PMC10257143
- DOI: 10.1126/scitranslmed.abn7336
BLIMP1 and NR4A3 transcription factors reciprocally regulate antitumor CAR T cell stemness and exhaustion
Abstract
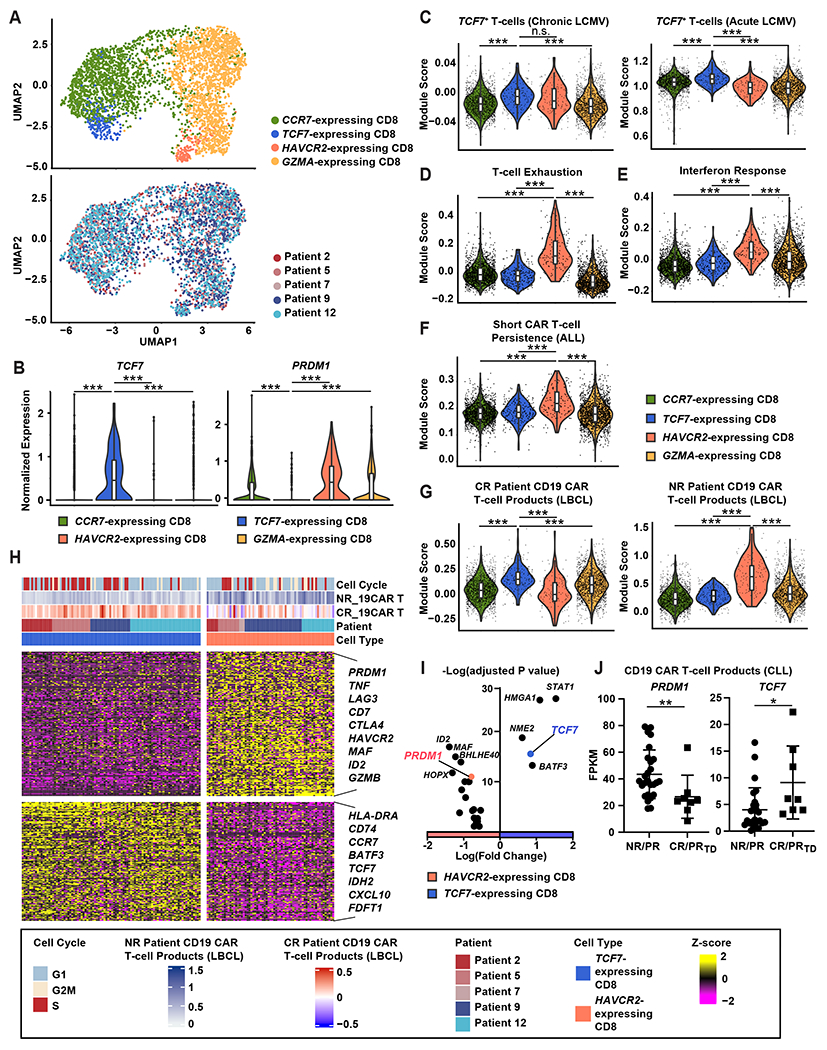
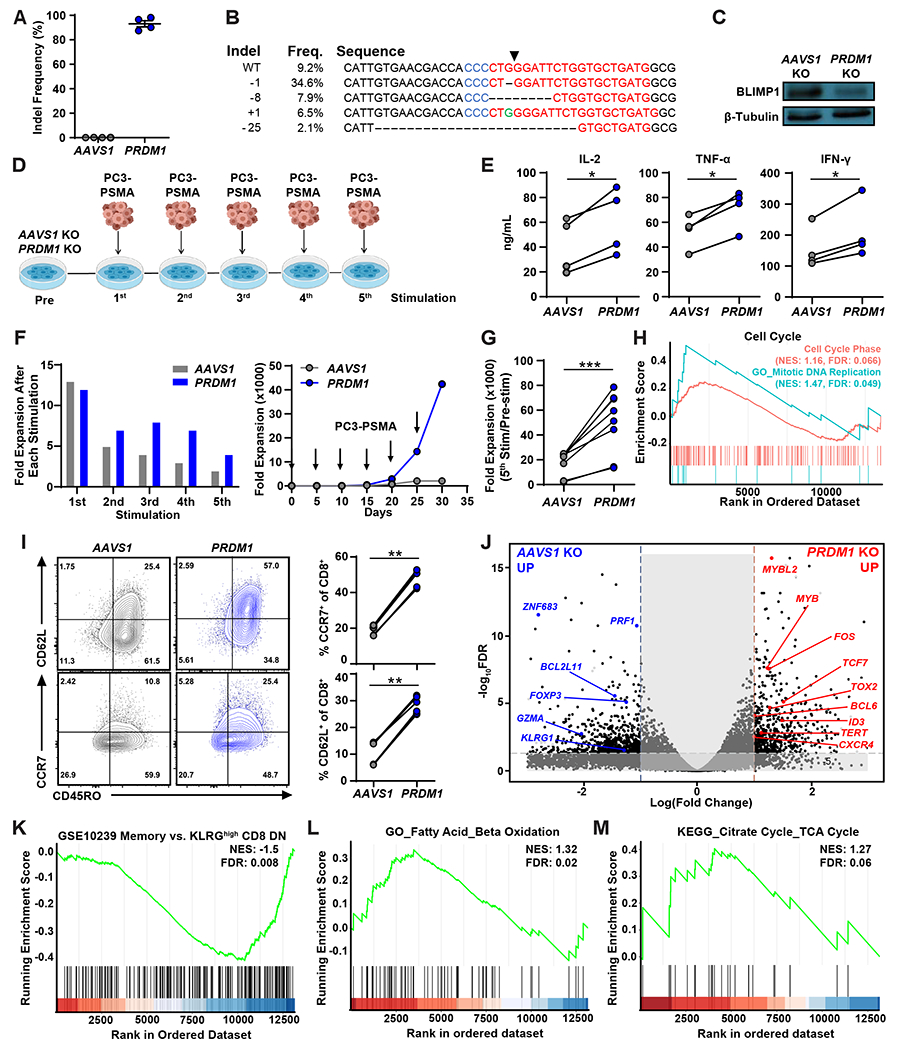
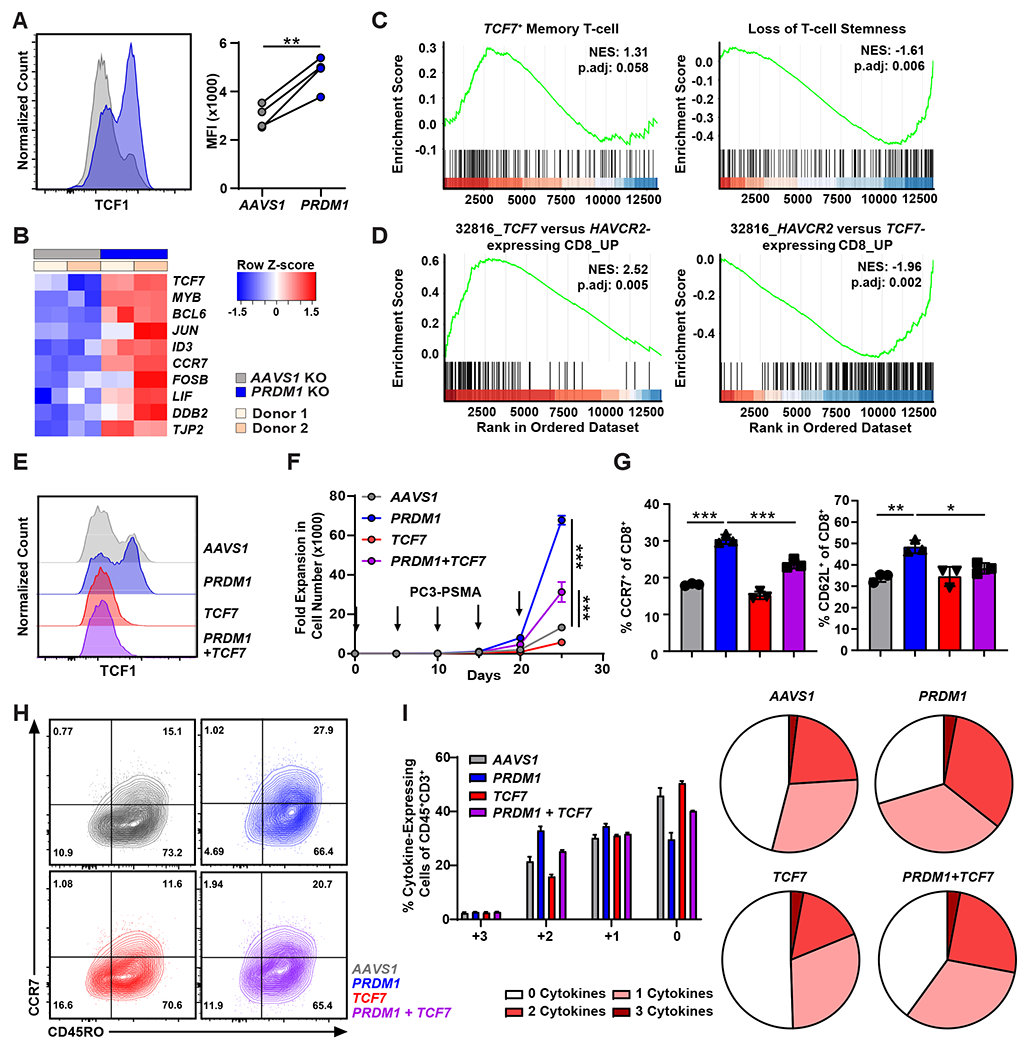
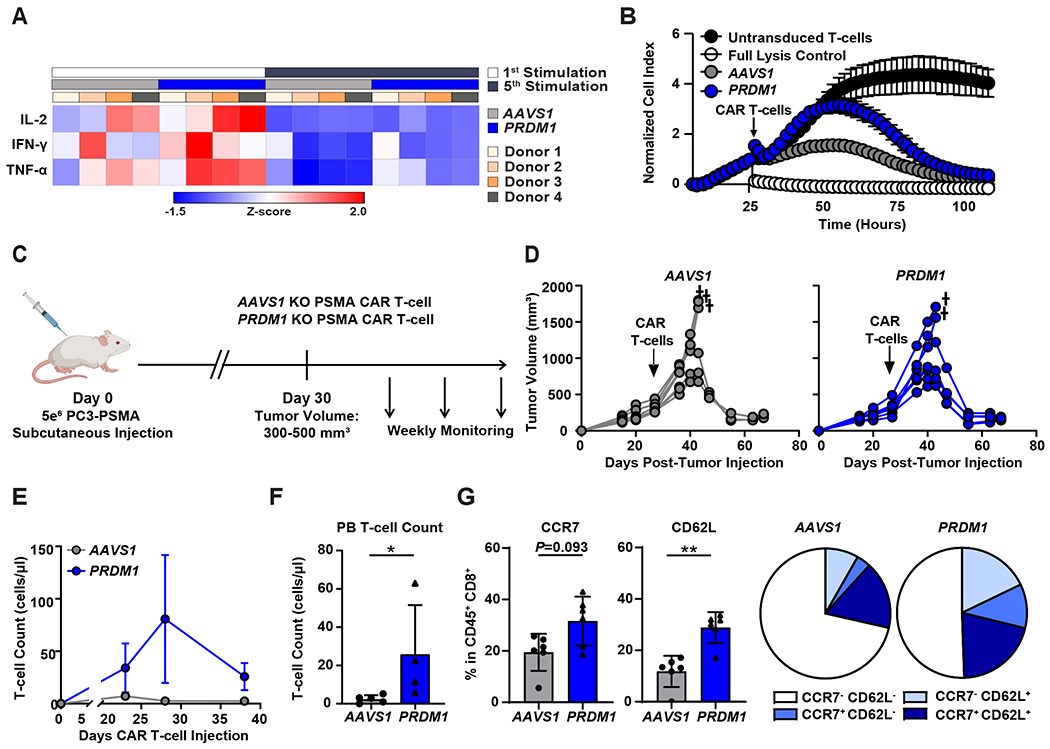
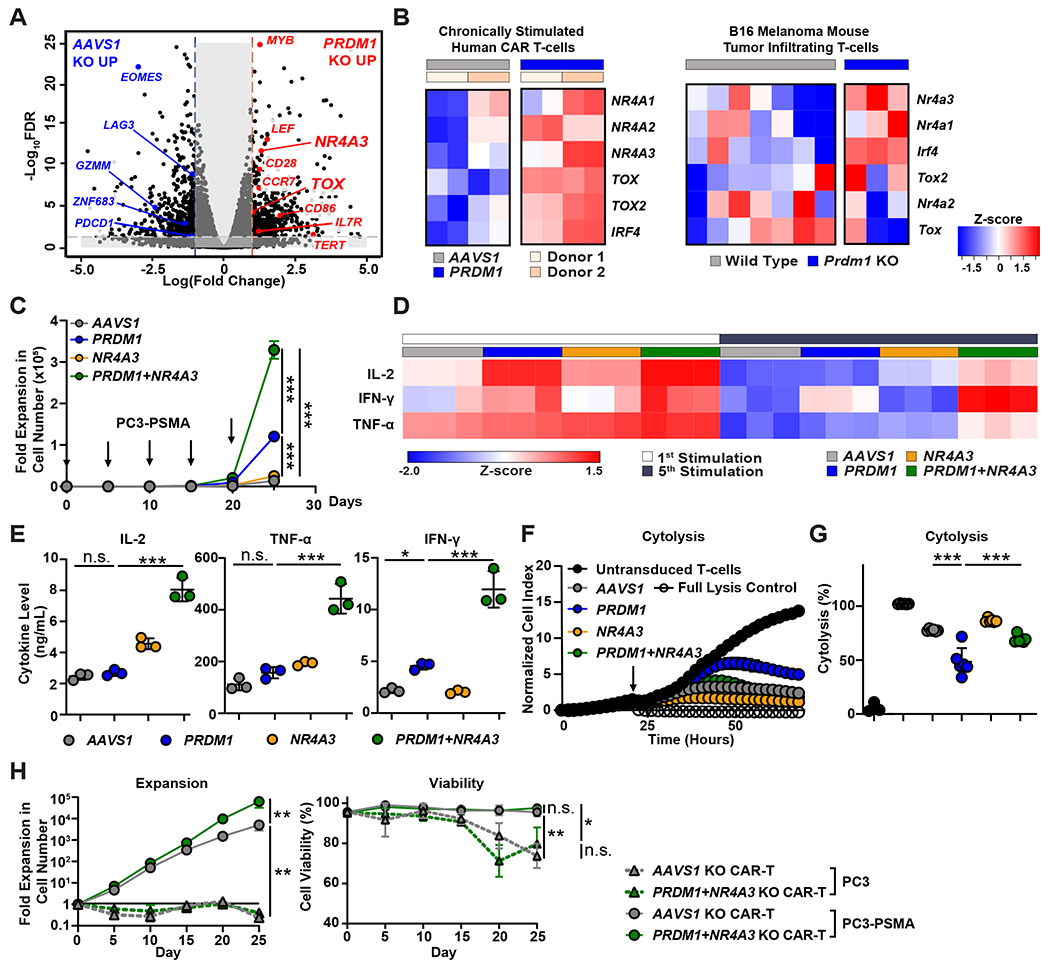
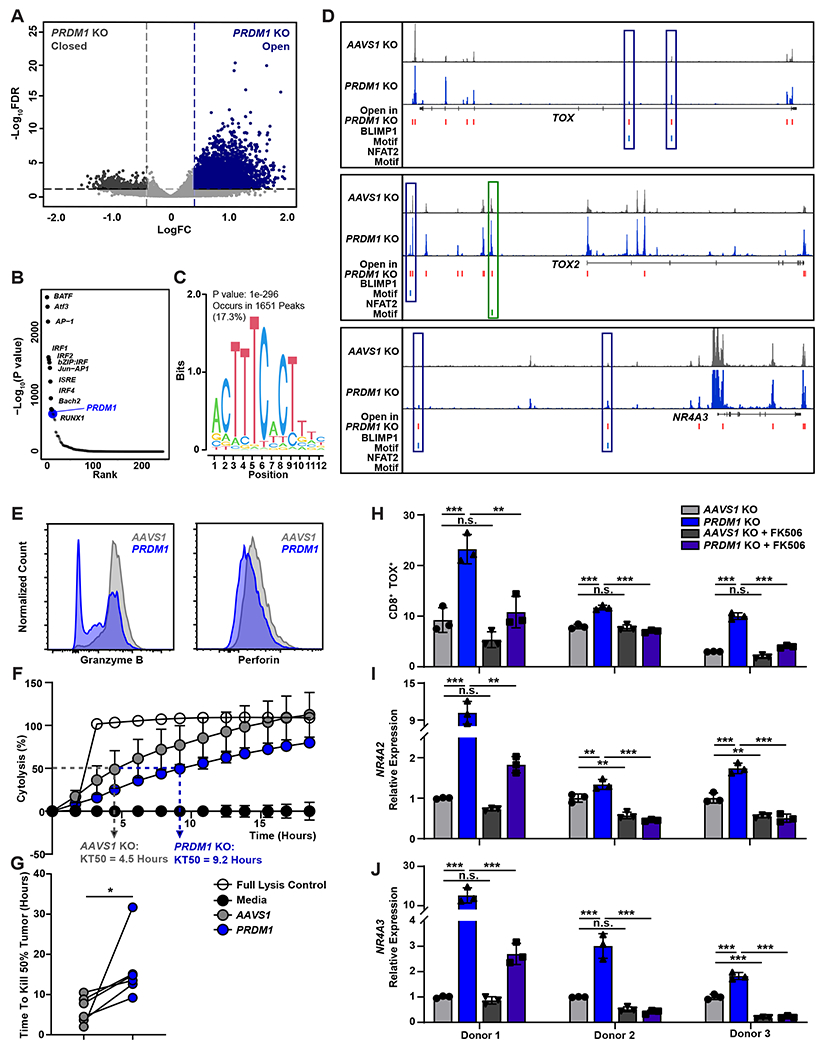
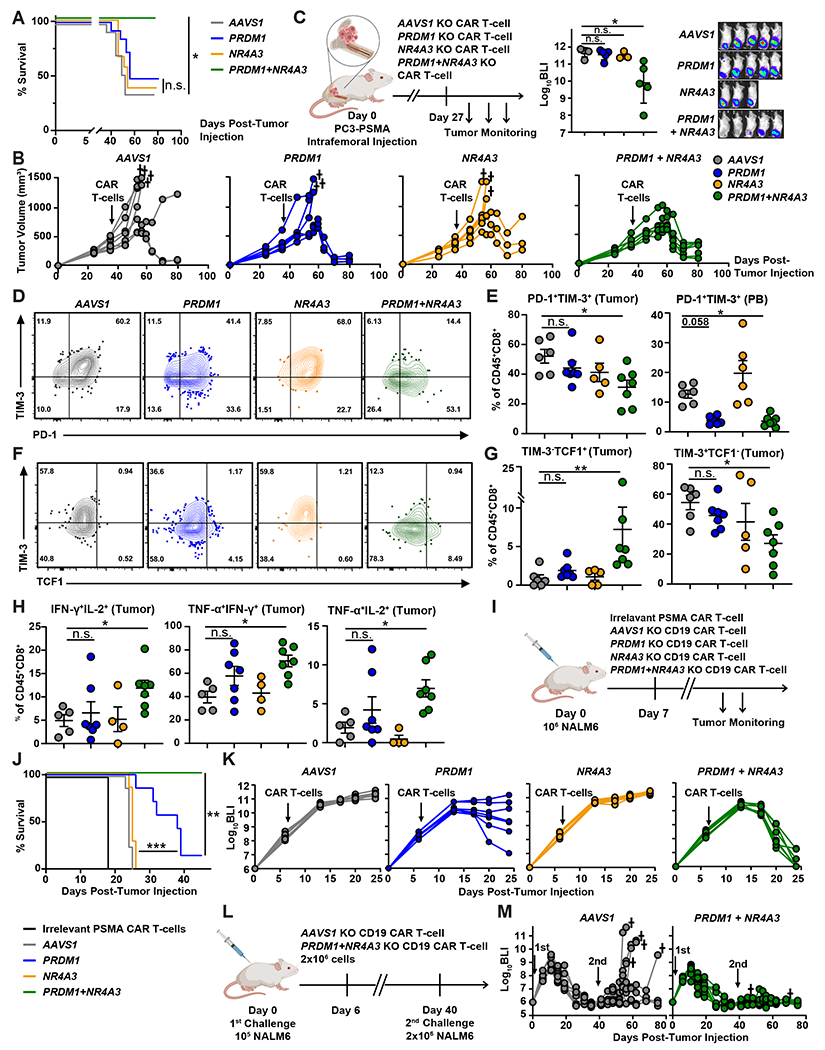
Chimeric antigen receptor (CAR) T cells have not induced meaningful clinical responses in solid tumors. Loss of T cell stemness, poor expansion capacity, and exhaustion during prolonged tumor antigen exposure are major causes of CAR T cell therapeutic resistance. Single-cell RNA-sequencing analysis of CAR T cells from a first-in-human trial in metastatic prostate cancer identified two independently validated cell states associated with antitumor potency or lack of efficacy. Low expression of PRDM1, encoding the BLIMP1 transcription factor, defined highly potent TCF7 [encoding T cell factor 1 (TCF1)]-expressing CD8+ CAR T cells, whereas enrichment of HAVCR2 [encoding T cell immunoglobulin and mucin-domain containing-3 (TIM-3)]-expressing CD8+ T cells with elevated PRDM1 was associated with poor outcomes. PRDM1 knockout promoted TCF7-dependent CAR T cell stemness and proliferation, resulting in marginally enhanced leukemia control in mice. However, in the setting of PRDM1 deficiency, a negative epigenetic feedback program of nuclear factor of activated T cells (NFAT)-driven T cell dysfunction was identified. This program was characterized by compensatory up-regulation of NR4A3 and other genes encoding exhaustion-related transcription factors that hampered T cell effector function in solid tumors. Dual knockout of PRDM1 and NR4A3 skewed CAR T cell phenotypes away from TIM-3+CD8+ and toward TCF1+CD8+ to counter exhaustion of tumor-infiltrating CAR T cells and improve antitumor responses, effects that were not achieved with PRDM1 and NR4A3 single knockout alone. These data underscore dual targeting of PRDM1 and NR4A3 as a promising approach to advance adoptive cell immuno-oncotherapy.
Conflict of interest statement
I.Y.J., S.F.L., M.M.D, D.L.S. and J.A.F. have filed patent applications in the field of T-cell therapy for cancer and have received royalties. Patents related to this work are as follows: Biomarkers and CAR T-cell therapies with enhanced efficacy (US20200087376A1) and CAR T-cell therapies with enhanced efficacy (US20210047405A1). M.M.D. and J.A.F. have received research funding from Tmunity Therapeutics. M.M.D. serves on the Scientific Advisory Board for Cellares Corporation. S.F.L. has served as a consultant for Novartis Pharmaceuticals, Kite Pharma, and Wugen and receives clinical trial funding from Novartis Pharmaceuticals. J.A.F. is a member of the Scientific Advisory Boards of Cartography Bio. and Shennon Biotechnologies Inc.
Figures
References
-
- Porter DL, Hwang W-T, Frey NV, Lacey SF, Shaw PA, Loren AW, Bagg A, Marcucci KT, Shen A, Gonzalez V, Ambrose D, Grupp SA, Chew A, Zheng Z, Milone MC, Levine BL, Melenhorst JJ, June CH, Chimeric antigen receptor T cells persist and induce sustained remissions in relapsed refractory chronic lymphocytic leukemia. Science Translational Medicine 7, 303ra139–303ra139 (2015); published online EpubSeptember 2, 2015 (10.1126/scitranslmed.aac5415). - DOI - PMC - PubMed
-
- Neelapu SS, Locke FL, Bartlett NL, Lekakis LJ, Miklos DB, Jacobson CA, Braunschweig I, Oluwole OO, Siddiqi T, Lin Y, Timmerman JM, Stiff PJ, Friedberg JW, Flinn IW, Goy A, Hill BT, Smith MR, Deol A, Farooq U, McSweeney P, Munoz J, Avivi I, Castro JE, Westin JR, Chavez JC, Ghobadi A, Komanduri KV, Levy R, Jacobsen ED, Witzig TE, Reagan P, Bot A, Rossi J, Navale L, Jiang Y, Aycock J, Elias M, Chang D, Wiezorek J, Go WY, Axicabtagene Ciloleucel CAR T-Cell Therapy in Refractory Large B-Cell Lymphoma. The New England Journal of Medicine 377, 2531–2544 (2017); published online Epub2017-12-28 (10.1056/NEJMoa1707447). - DOI - PMC - PubMed
-
- Maude SL, Laetsch TW, Buechner J, Rives S, Boyer M, Bittencourt H, Bader P, Verneris MR, Stefanski HE, Myers GD, Qayed M, De Moerloose B, Hiramatsu H, Schlis K, Davis KL, Martin PL, Nemecek ER, Yanik GA, Peters C, Baruchel A, Boissel N, Mechinaud F, Balduzzi A, Krueger J, June CH, Levine BL, Wood P, Taran T, Leung M, Mueller KT, Zhang Y, Sen K, Lebwohl D, Pulsipher MA, Grupp SA, Tisagenlecleucel in Children and Young Adults with B-Cell Lymphoblastic Leukemia. The New England Journal of Medicine 378, 439–448 (2018); published online Epub2018-02-01 (10.1056/NEJMoa1709866). - DOI - PMC - PubMed
-
- Raje N, Berdeja J, Lin Y, Siegel D, Jagannath S, Madduri D, Liedtke M, Rosenblatt J, Maus MV, Turka A, Lam LP, Morgan RA, Friedman K, Massaro M, Wang J, Russotti G, Yang Z, Campbell T, Hege K, Petrocca F, Quigley MT, Munshi N, Kochenderfer JN, Anti-BCMA CAR T-Cell Therapy bb2121 in Relapsed or Refractory Multiple Myeloma. N Engl J Med 380, 1726–1737 (2019); published online EpubMay 2 (10.1056/NEJMoa1817226). - DOI - PMC - PubMed
-
- Cohen AD, Garfall AL, Stadtmauer EA, Melenhorst JJ, Lacey SF, Lancaster E, Vogl DT, Weiss BM, Dengel K, Nelson A, Plesa G, Chen F, Davis MM, Hwang WT, Young RM, Brogdon JL, Isaacs R, Pruteanu-Malinici I, Siegel DL, Levine BL, June CH, Milone MC, B cell maturation antigen-specific CAR T cells are clinically active in multiple myeloma. J Clin Invest 129, 2210–2221 (2019); published online EpubMar 21 (10.1172/JCI126397). - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
