

A high-quality, haplotype-phased genome reconstruction reveals unexpected haplotype diversity in a pearl oyster
- PMID: 36351462
- PMCID: PMC9646362
- DOI: 10.1093/dnares/dsac035
A high-quality, haplotype-phased genome reconstruction reveals unexpected haplotype diversity in a pearl oyster
Abstract
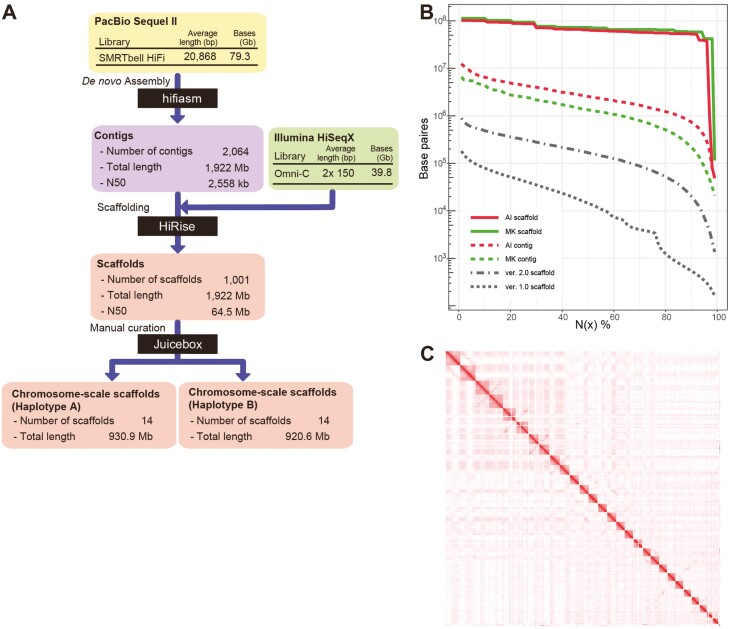
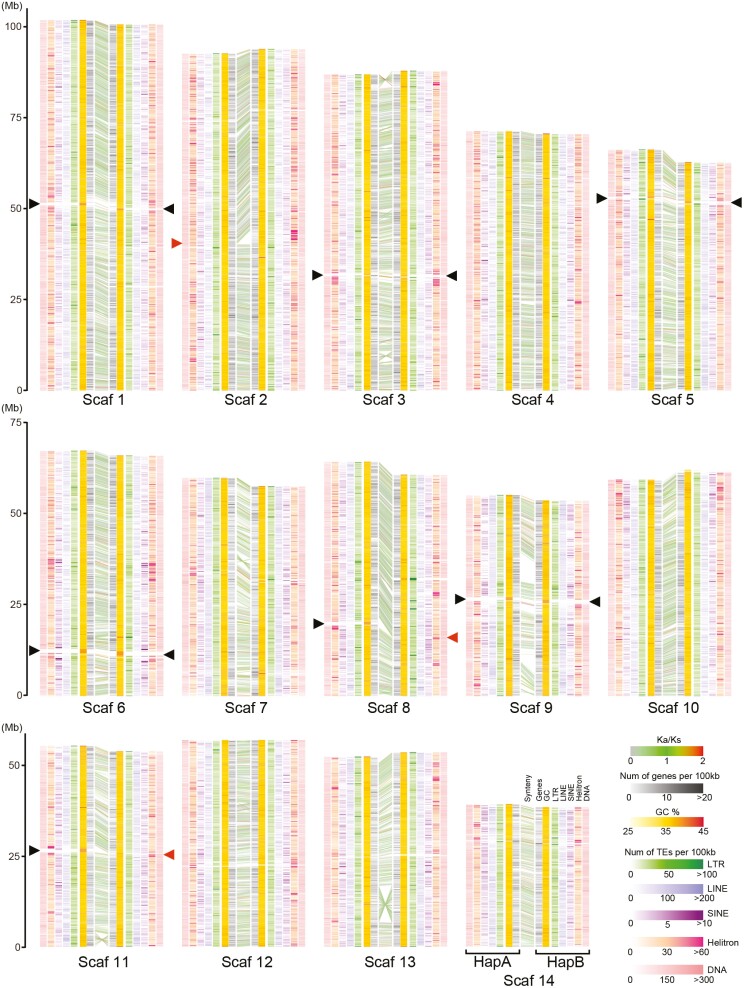
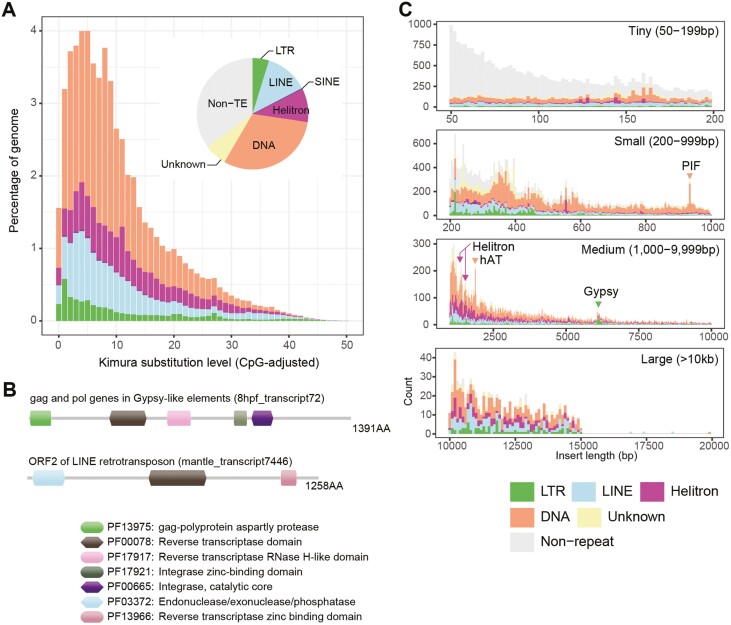
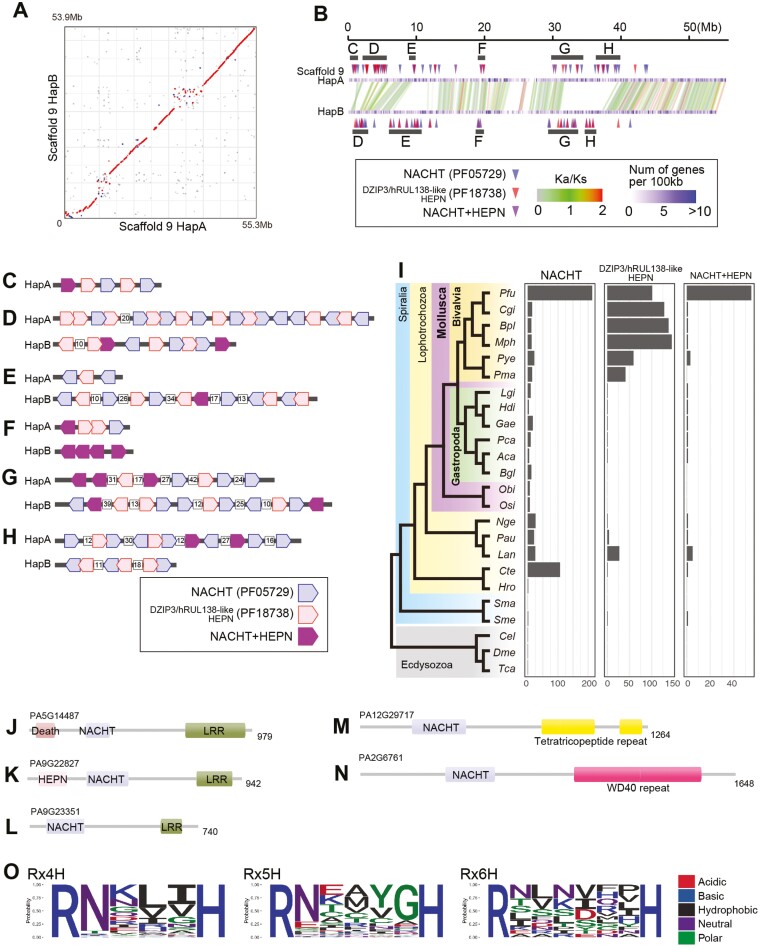
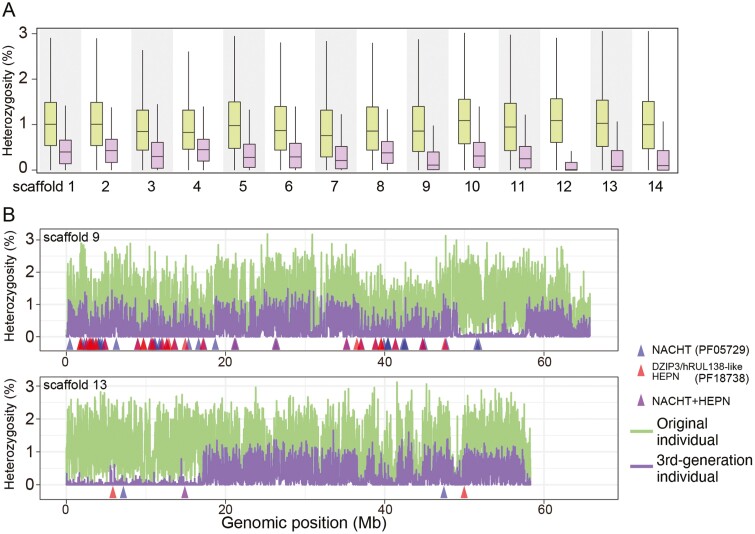
Homologous chromosomes in the diploid genome are thought to contain equivalent genetic information, but this common concept has not been fully verified in animal genomes with high heterozygosity. Here we report a near-complete, haplotype-phased, genome assembly of the pearl oyster, Pinctada fucata, using hi-fidelity (HiFi) long reads and chromosome conformation capture data. This assembly includes 14 pairs of long scaffolds (>38 Mb) corresponding to chromosomes (2n = 28). The accuracy of the assembly, as measured by an analysis of k-mers, is estimated to be 99.99997%. Moreover, the haplotypes contain 95.2% and 95.9%, respectively, complete and single-copy BUSCO genes, demonstrating the high quality of the assembly. Transposons comprise 53.3% of the assembly and are a major contributor to structural variations. Despite overall collinearity between haplotypes, one of the chromosomal scaffolds contains megabase-scale non-syntenic regions, which necessarily have never been detected and resolved in conventional haplotype-merged assemblies. These regions encode expanded gene families of NACHT, DZIP3/hRUL138-like HEPN, and immunoglobulin domains, multiplying the immunity gene repertoire, which we hypothesize is important for the innate immune capability of pearl oysters. The pearl oyster genome provides insight into remarkable haplotype diversity in animals.
Keywords: Mollusca; aquaculture; haplotype-phased genome assembly; immunity; pearl oyster.
© The Author(s) 2022. Published by Oxford University Press on behalf of Kazusa DNA Research Institute.
Figures
Similar articles
-
A chromosome-level, haplotype-resolved genome assembly and annotation for the Eurasian minnow (Leuciscidae: Phoxinus phoxinus) provide evidence of haplotype diversity.Gigascience. 2025 Jan 6;14:giae116. doi: 10.1093/gigascience/giae116. Gigascience. 2025. PMID: 39877992 Free PMC article.
-
The haplotype-resolved chromosome pairs of a heterozygous diploid African cassava cultivar reveal novel pan-genome and allele-specific transcriptome features.Gigascience. 2022 Mar 24;11:giac028. doi: 10.1093/gigascience/giac028. Gigascience. 2022. PMID: 35333302 Free PMC article.
-
Draft genome of the pearl oyster Pinctada fucata: a platform for understanding bivalve biology.DNA Res. 2012 Apr;19(2):117-30. doi: 10.1093/dnares/dss005. Epub 2012 Feb 6. DNA Res. 2012. PMID: 22315334 Free PMC article.
-
Bivalve-specific gene expansion in the pearl oyster genome: implications of adaptation to a sessile lifestyle.Zoological Lett. 2016 Feb 18;2:3. doi: 10.1186/s40851-016-0039-2. eCollection 2016. Zoological Lett. 2016. PMID: 26900483 Free PMC article.
-
The importance of total genome databases in research on Akoya pearl oyster.Zoolog Sci. 2013 Oct;30(10):781-2. doi: 10.2108/zsj.30.781. Zoolog Sci. 2013. PMID: 24125641 Review.
Cited by
-
Chromosome-level genome assembly of Pontederia cordata L. provides insights into its rapid adaptation and variation of flower colours.DNA Res. 2025 Mar 1;32(2):dsaf002. doi: 10.1093/dnares/dsaf002. DNA Res. 2025. PMID: 39878035 Free PMC article.
-
The chromosome level genome assembly of the Asian green mussel, Perna viridis.Sci Data. 2024 Aug 28;11(1):930. doi: 10.1038/s41597-024-03802-2. Sci Data. 2024. PMID: 39198463 Free PMC article.
-
Multi-omic insights into the formation and evolution of a novel shell microstructure in oysters.BMC Biol. 2023 Sep 29;21(1):204. doi: 10.1186/s12915-023-01706-y. BMC Biol. 2023. PMID: 37775818 Free PMC article.
-
A chromosome-level, haplotype-resolved genome assembly and annotation for the Eurasian minnow (Leuciscidae: Phoxinus phoxinus) provide evidence of haplotype diversity.Gigascience. 2025 Jan 6;14:giae116. doi: 10.1093/gigascience/giae116. Gigascience. 2025. PMID: 39877992 Free PMC article.
-
The First High-Quality Genome Assembly of Freshwater Pearl Mussel Sinohyriopsis cumingii: New Insights into Pearl Biomineralization.Int J Mol Sci. 2024 Mar 9;25(6):3146. doi: 10.3390/ijms25063146. Int J Mol Sci. 2024. PMID: 38542120 Free PMC article.
References
MeSH terms
LinkOut - more resources
Full Text Sources
Miscellaneous
