

Co-immunization with hemagglutinin stem immunogens elicits cross-group neutralizing antibodies and broad protection against influenza A viruses
- PMID: 36356572
- PMCID: PMC9772109
- DOI: 10.1016/j.immuni.2022.10.015
Co-immunization with hemagglutinin stem immunogens elicits cross-group neutralizing antibodies and broad protection against influenza A viruses
Abstract
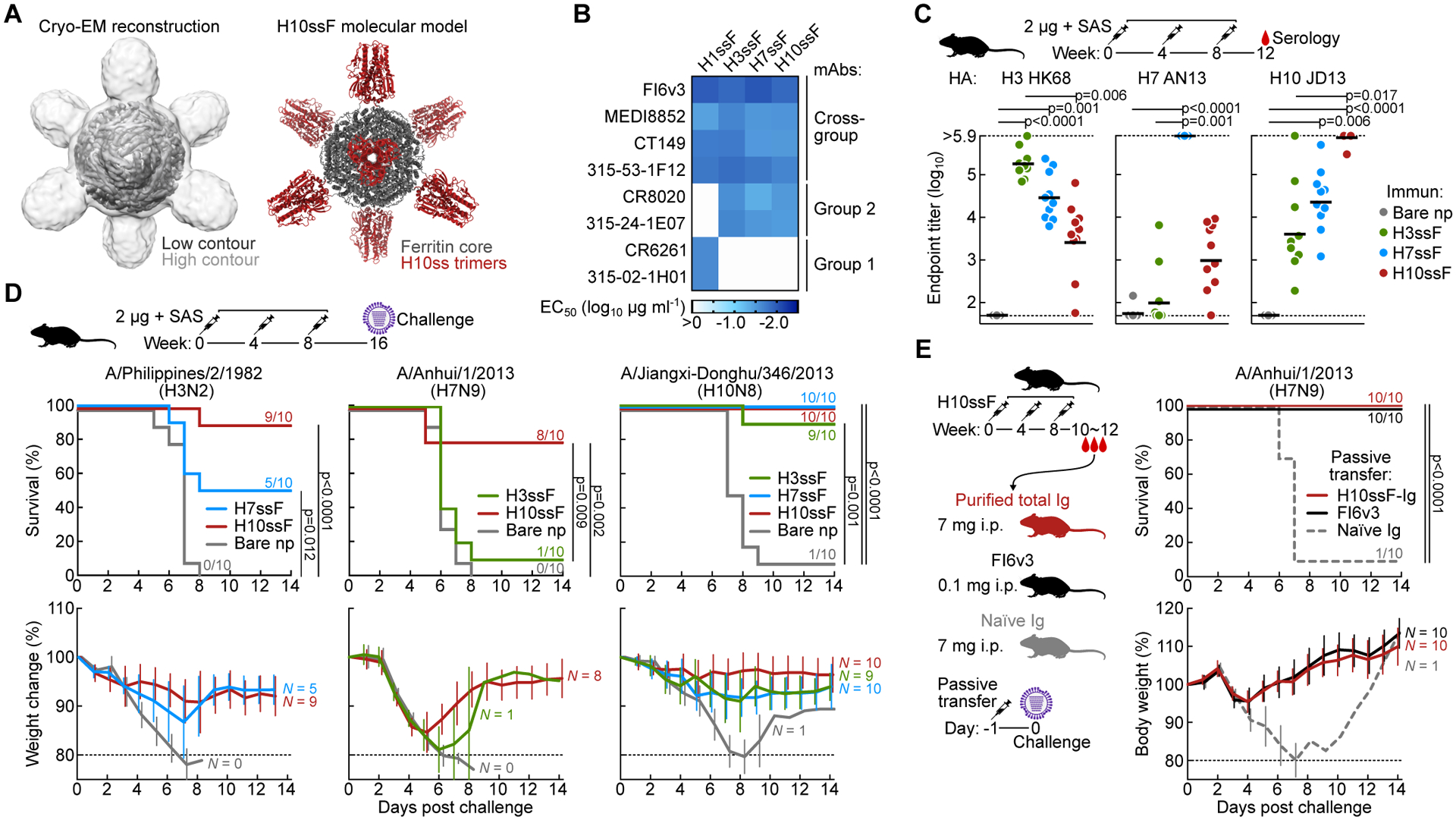
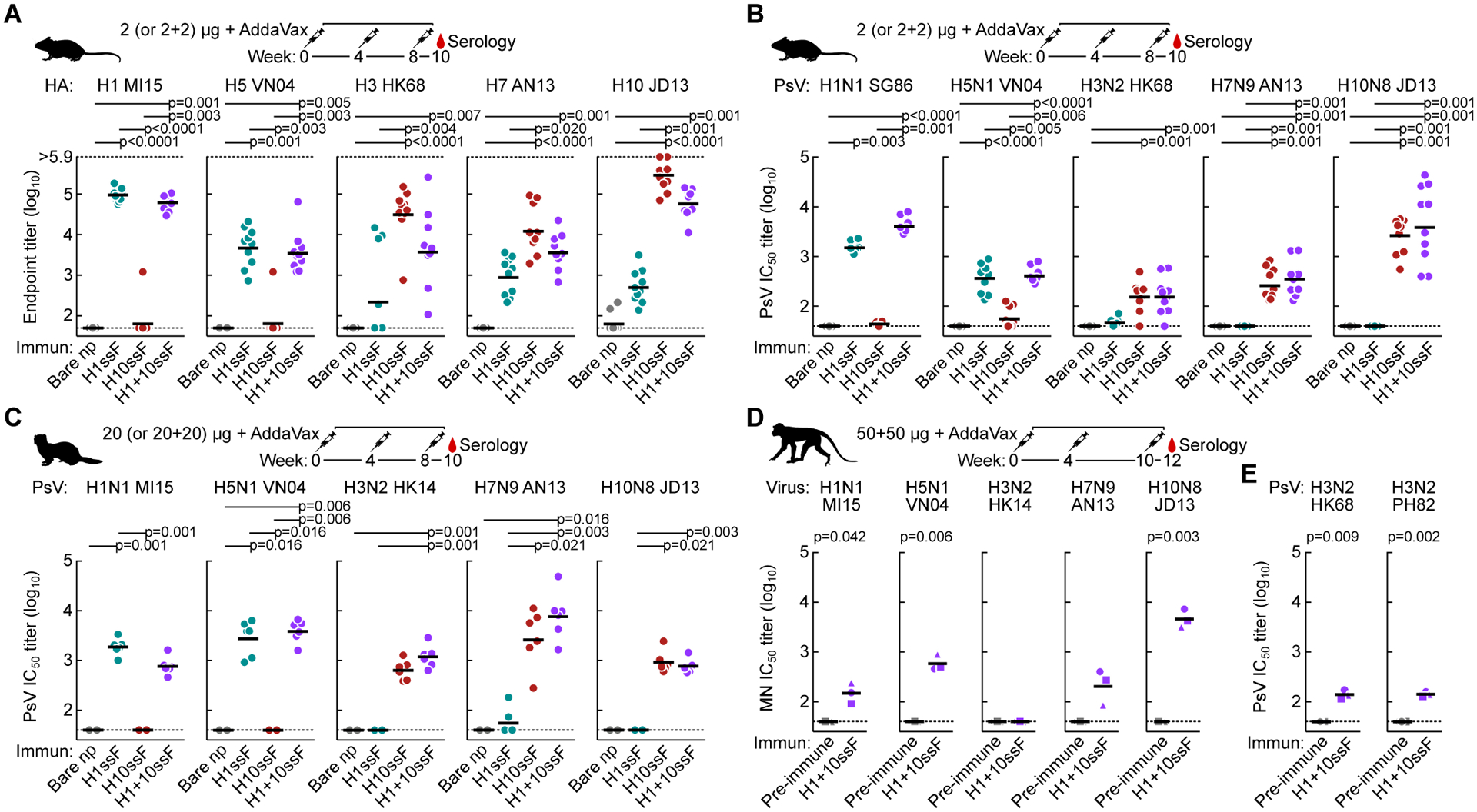
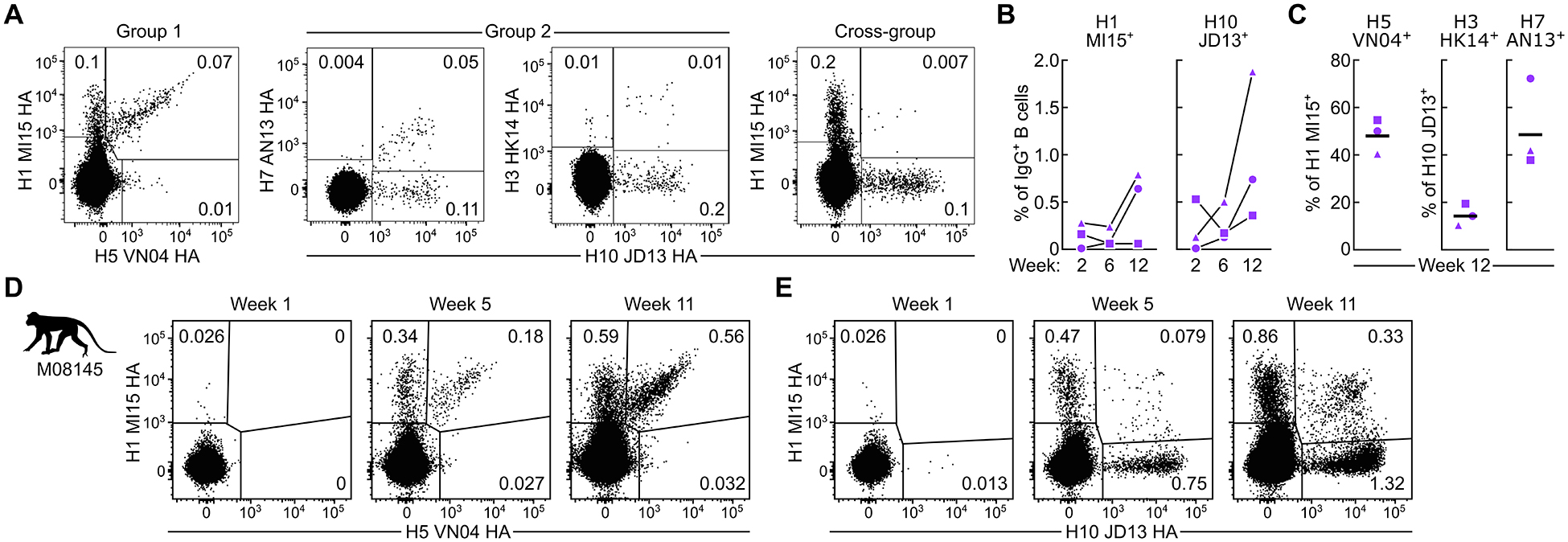
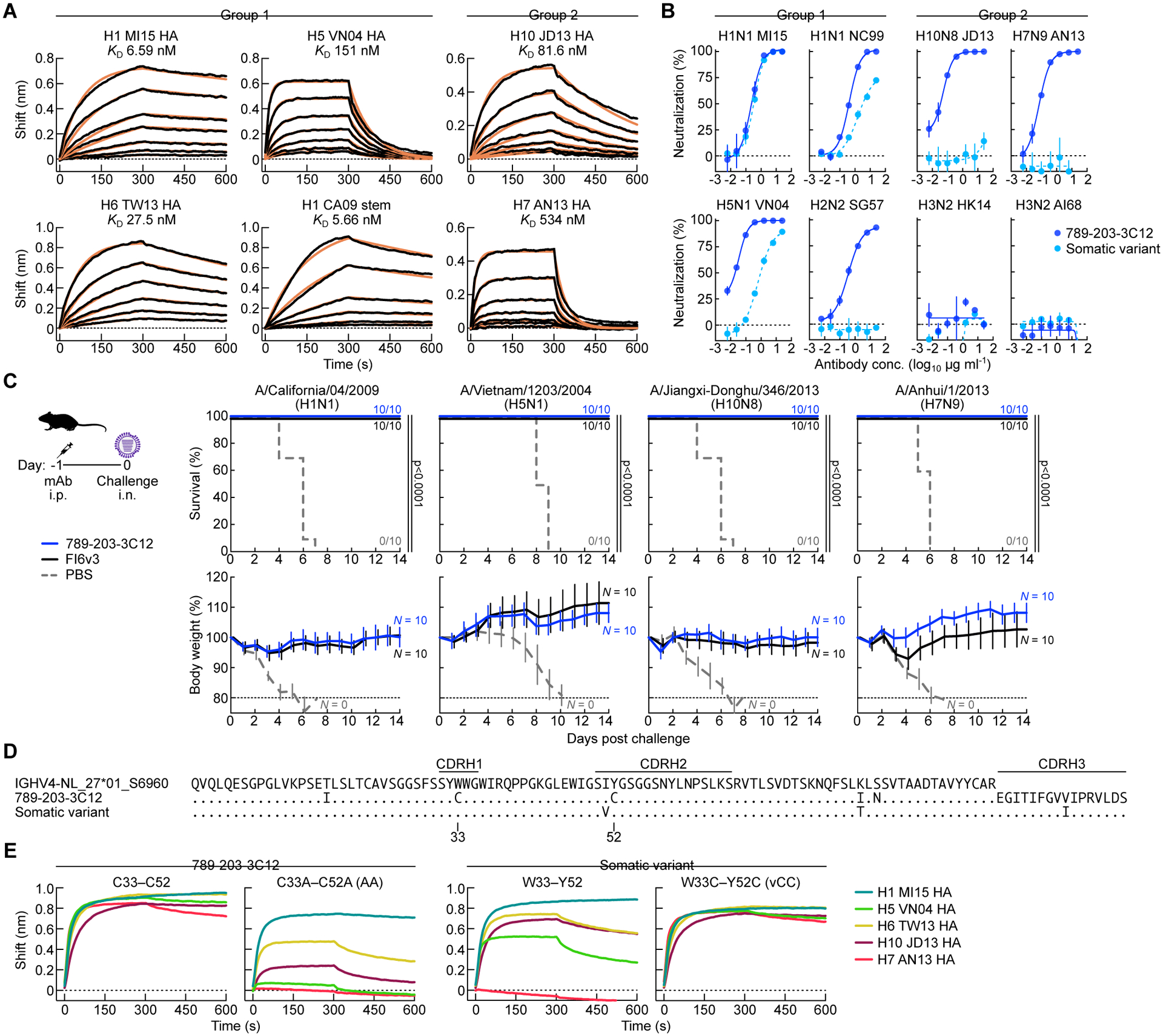
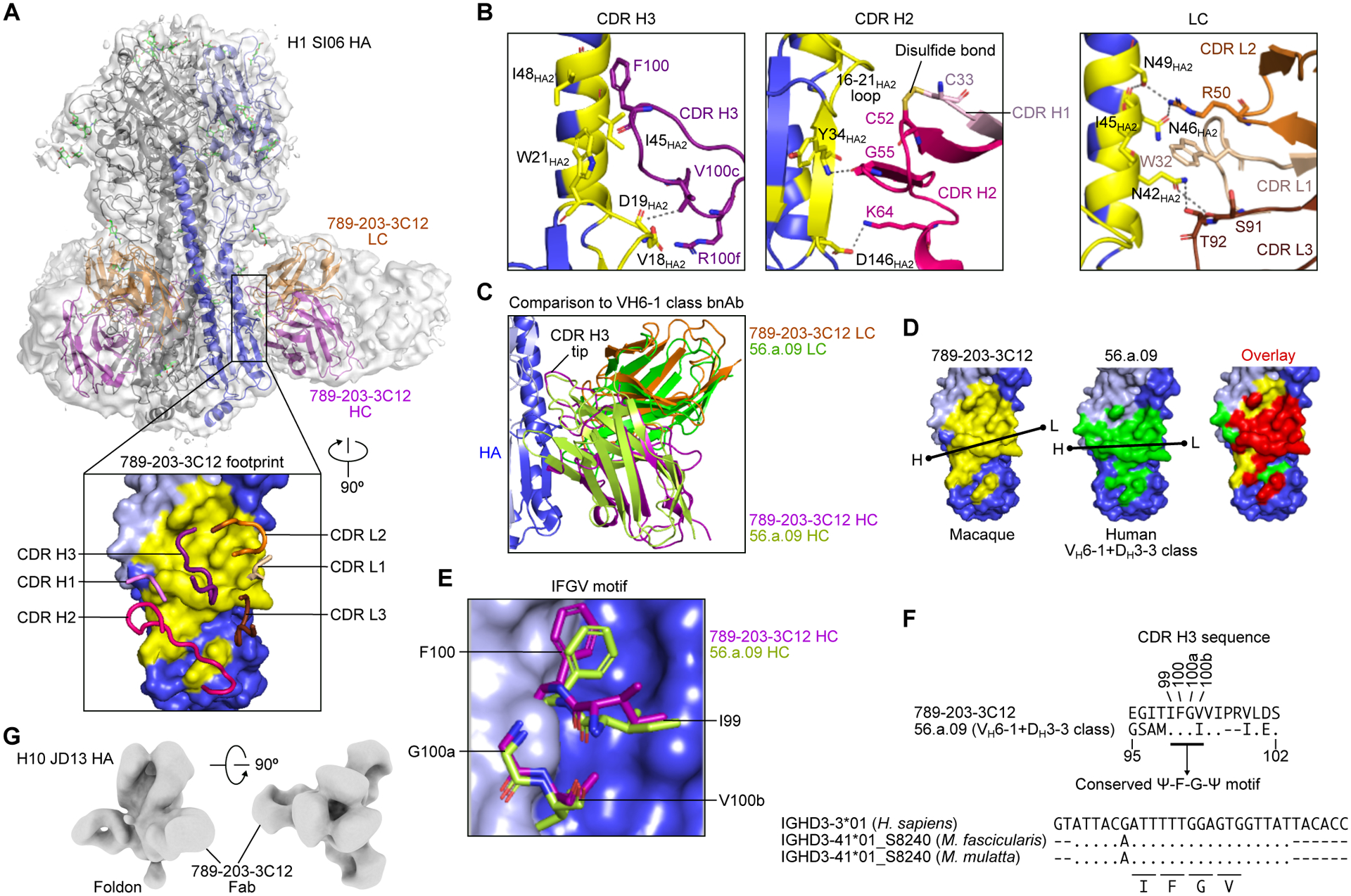
Current influenza vaccines predominantly induce immunity to the hypervariable hemagglutinin (HA) head, requiring frequent vaccine reformulation. Conversely, the immunosubdominant yet conserved HA stem harbors a supersite that is targeted by broadly neutralizing antibodies (bnAbs), representing a prime target for universal vaccines. Here, we showed that the co-immunization of two HA stem immunogens derived from group 1 and 2 influenza A viruses elicits cross-group protective immunity and neutralizing antibody responses in mice, ferrets, and nonhuman primates (NHPs). Immunized mice were protected from multiple group 1 and 2 viruses, and all animal models showed broad serum-neutralizing activity. A bnAb isolated from an immunized NHP broadly neutralized and protected against diverse viruses, including H5N1 and H7N9. Genetic and structural analyses revealed strong homology between macaque and human bnAbs, illustrating common biophysical constraints for acquiring cross-group specificity. Vaccine elicitation of stem-directed cross-group-protective immunity represents a step toward the development of broadly protective influenza vaccines.
Keywords: HA stalk; HA stem; bnAb; cryo-EM; ferritin; influenza; nanoparticle; nonhuman primate; pandemic preparedness; vaccine.
Published by Elsevier Inc.
Conflict of interest statement
Declaration of interests S.M.M., J.C.B., P.D.K., J.R.M., B.S.G., and M.K. are named inventors of US patents 9,441,019, 10,137,190, 10,363,301, and 11,338,033 on influenza HA nanoparticle vaccines and stabilized HA stem trimers and of several pending applications on related technologies filed by the US Department of Health and Human Services (NIH).
Figures
References
-
- Andrews SF, Joyce MG, Chambers MJ, Gillespie RA, Kanekiyo M, Leung K, Yang ES, Tsybovsky Y, Wheatley AK, Crank MC, et al. (2017). Preferential induction of cross-group influenza A hemagglutinin stem-specific memory B cells after H7N9 immunization in humans. Sci Immunol 2, eaan2676. 10.1126/sciimmunol.aan2676. - DOI - PubMed
-
- Avnir Y, Watson CT, Glanville J, Peterson EC, Tallarico AS, Bennett AS, Qin K, Fu Y, Huang C-Y, Beigel JH, et al. (2016). IGHV1–69 polymorphism modulates anti-influenza antibody repertoires, correlates with IGHV utilization shifts and varies by ethnicity. Sci. Rep 6, 20842. 10.1038/srep20842. - DOI - PMC - PubMed
-
- Bernstein DI, Guptill J, Naficy A, Nachbagauer R, Berlanda-Scorza F, Feser J, Wilson PC, Solórzano A, Van der Wielen M, Walter EB, et al. (2020). Immunogenicity of chimeric haemagglutinin-based, universal influenza virus vaccine candidates: interim results of a randomised, placebo-controlled, phase 1 clinical trial. Lancet Infect. Dis 20, 80–91. 10.1016/S1473-3099(19)30393-7. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
