

Proteomic Profiling Revealed Mitochondrial Dysfunction in Photoreceptor Cells under Hyperglycemia
- PMID: 36362154
- PMCID: PMC9658613
- DOI: 10.3390/ijms232113366
Proteomic Profiling Revealed Mitochondrial Dysfunction in Photoreceptor Cells under Hyperglycemia
Abstract
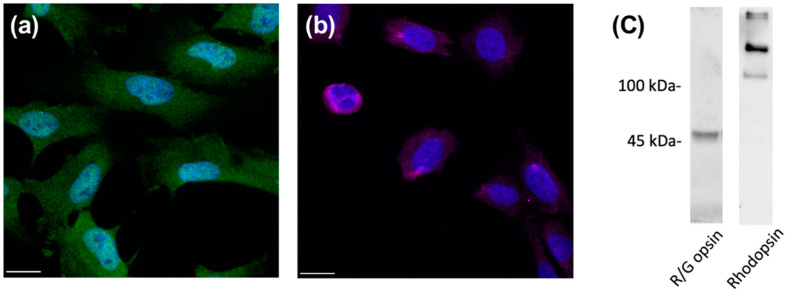
Diabetic retinopathy (DR) was identified as a leading cause of blindness and vision impairment in 2020. In addition to vasculopathy, DR has been found to involve retinal neurons, including amacrine cells and retinal ganglion cells. Despite possessing features that are susceptible to diabetic conditions, photoreceptor cells have received relatively little attention with respect to the development of DR. Until recently, studies have suggested that photoreceptors secret proinflammatory molecules and produce reactive oxygen species that contribute to the development of DR. However, the effect of hyperglycemia on photoreceptors and its underlying mechanism remains elusive. In this study, the direct effect of high glucose on photoreceptor cells was investigated using a 661w photoreceptor-like cell line. A data-independent sequential window acquisition of all theoretical mass spectra (SWATH)-based proteomic approach was employed to study changes induced by high glucose in the proteomic profile of the cells. The results indicated that high glucose induced a significant increase in apoptosis and ROS levels in the 661w cells, with mitochondrial dysfunction among the major affected canonical pathways. The involvement of mitochondrial dysfunction was further supported by increased mitochondrial fission and reduced mitochondrial bioenergetics. Collectively, these findings provide a biological basis for a possible role of photoreceptors in the pathogenesis of DR.
Keywords: SWATH-MS; diabetic retinopathy; mitochondrial dysfunction.
Conflict of interest statement
The authors declare no conflict of interest.
Figures







Similar articles
-
Uncovering the Therapeutic Potential of Phosphocreatine in Diabetic Retinopathy: Mitigating Mitochondrial Dysfunction and Apoptosis via JAK2/STAT3 Signaling Pathway.J Mol Neurosci. 2024 Jan 17;74(1):11. doi: 10.1007/s12031-023-02175-2. J Mol Neurosci. 2024. PMID: 38231435
-
Somatostatin protects photoreceptor cells against high glucose-induced apoptosis.Mol Vis. 2016 Dec 30;22:1522-1531. eCollection 2016. Mol Vis. 2016. PMID: 28050125 Free PMC article.
-
Glucose-mediated de novo lipogenesis in photoreceptors drives early diabetic retinopathy.J Biol Chem. 2021 Sep;297(3):101104. doi: 10.1016/j.jbc.2021.101104. Epub 2021 Aug 20. J Biol Chem. 2021. PMID: 34425110 Free PMC article.
-
Hypoxia and oxidative stress in the causation of diabetic retinopathy.Curr Diabetes Rev. 2011 Sep;7(5):291-304. doi: 10.2174/157339911797415620. Curr Diabetes Rev. 2011. PMID: 21916837 Review.
-
Diabetic retinopathy pathogenesis and the ameliorating effects of melatonin; involvement of autophagy, inflammation and oxidative stress.Life Sci. 2018 Jan 15;193:20-33. doi: 10.1016/j.lfs.2017.12.001. Epub 2017 Dec 5. Life Sci. 2018. PMID: 29203148 Review.
Cited by
-
Functional and structural changes in the neuroretina are accompanied by mitochondrial dysfunction in a type 2 diabetic mouse model.Eye Vis (Lond). 2023 Sep 1;10(1):37. doi: 10.1186/s40662-023-00353-2. Eye Vis (Lond). 2023. PMID: 37653465 Free PMC article.
-
Posttranslational modifications of proteins in diseased retina.Front Cell Neurosci. 2023 Mar 30;17:1150220. doi: 10.3389/fncel.2023.1150220. eCollection 2023. Front Cell Neurosci. 2023. PMID: 37066080 Free PMC article. Review.
-
Exploring Monkeypox: prospects for therapeutics through computational-aided drug discovery.Mol Divers. 2024 Oct;28(5):3497-3521. doi: 10.1007/s11030-023-10767-8. Epub 2023 Dec 11. Mol Divers. 2024. PMID: 38079063 Review.
-
Heyingwuzi formulation alleviates diabetic retinopathy by promoting mitophagy via the HIF-1α/BNIP3/NIX axis.World J Diabetes. 2024 Jun 15;15(6):1317-1339. doi: 10.4239/wjd.v15.i6.1317. World J Diabetes. 2024. PMID: 38983802 Free PMC article.
-
Coenzyme Q10 eyedrops conjugated with vitamin E TPGS alleviate neurodegeneration and mitochondrial dysfunction in the diabetic mouse retina.Front Cell Neurosci. 2024 May 28;18:1404987. doi: 10.3389/fncel.2024.1404987. eCollection 2024. Front Cell Neurosci. 2024. PMID: 38863499 Free PMC article.
References
-
- Leasher J.L., Bourne R.R., Flaxman S.R., Jonas J.B., Keeffe J., Naidoo K., Pesudovs K., Price H., White R.A., Wong T.Y. Global estimates on the number of people blind or visually impaired by diabetic retinopathy: A meta-analysis from 1990 to 2010. Diabetes Care. 2016;39:1643–1649. doi: 10.2337/dc15-2171. - DOI - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
