

Host-Directed Therapies for Tuberculosis
- PMID: 36365041
- PMCID: PMC9697779
- DOI: 10.3390/pathogens11111291
Host-Directed Therapies for Tuberculosis
Abstract
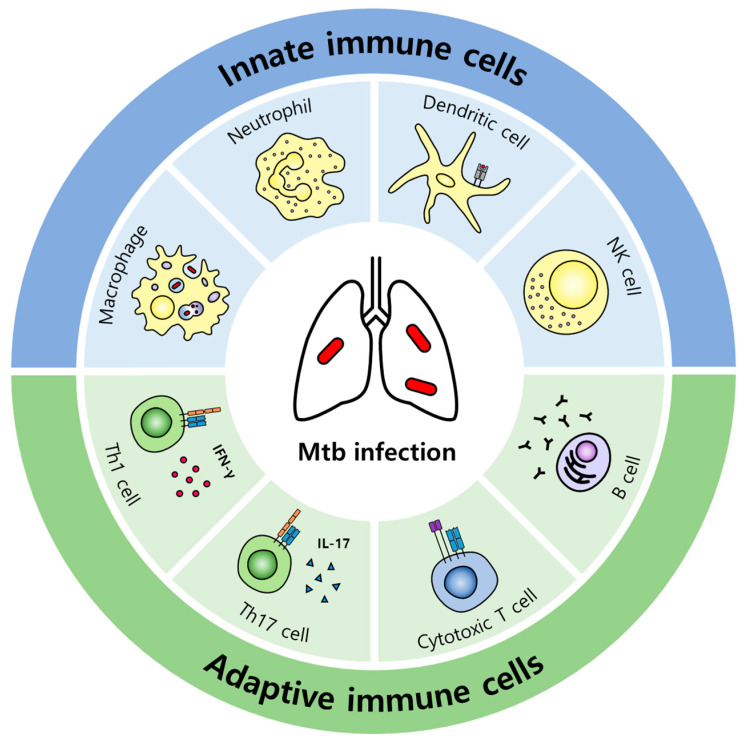
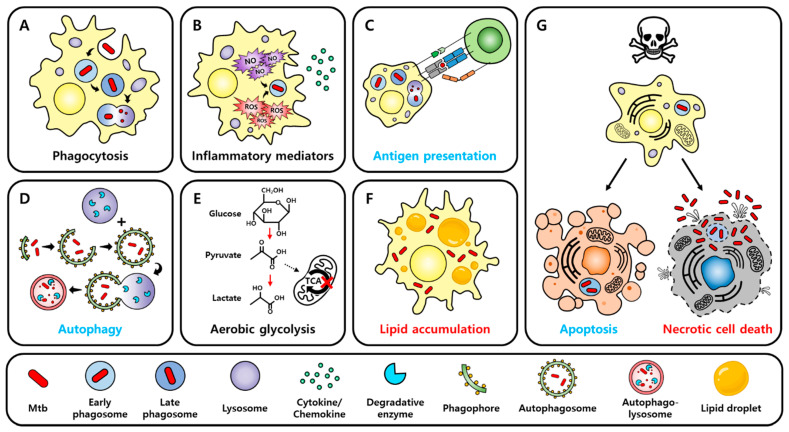
Tuberculosis (TB) is one of the leading causes of death worldwide, consistently threatening public health. Conventional tuberculosis treatment requires a long-term treatment regimen and is associated with side effects. The efficacy of antitubercular drugs has decreased with the emergence of drug-resistant TB; therefore, the development of new TB treatment strategies is urgently needed. In this context, we present host-directed therapy (HDT) as an alternative to current tuberculosis therapy. Unlike antitubercular drugs that directly target Mycobacterium tuberculosis (Mtb), the causative agent of TB, HDT is an approach for treating TB that appropriately modulates host immune responses. HDT primarily aims to enhance the antimicrobial activity of the host in order to control Mtb infection and attenuate excessive inflammation in order to minimize tissue damage. Recently, research based on the repositioning of drugs for use in HDT has been in progress. Based on the overall immune responses against Mtb infection and the immune-evasion mechanisms of Mtb, this review examines the repositioned drugs available for HDT and their mechanisms of action.
Keywords: Mycobacterium tuberculosis; drug repositioning; host-directed therapy; tuberculosis.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
