

The basolateral amygdala to lateral septum circuit is critical for regulating social novelty in mice
- PMID: 36369482
- PMCID: PMC9852457
- DOI: 10.1038/s41386-022-01487-y
The basolateral amygdala to lateral septum circuit is critical for regulating social novelty in mice
Abstract
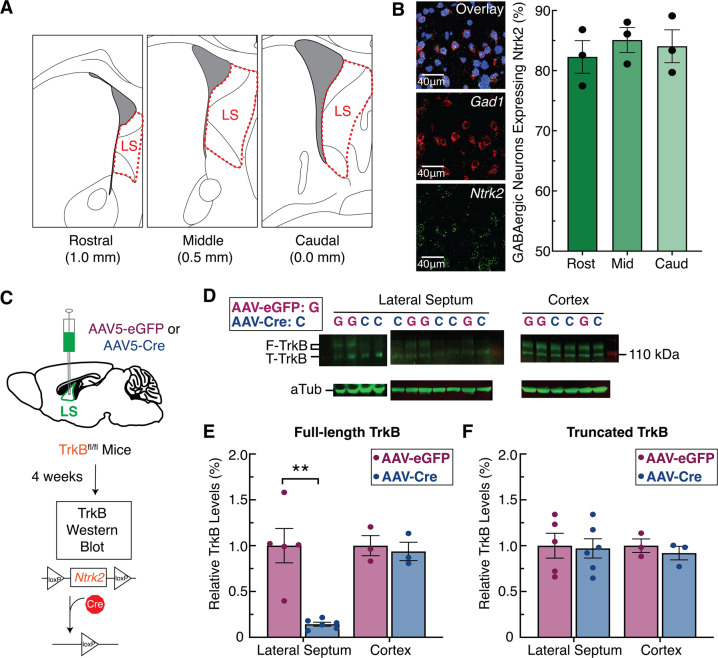
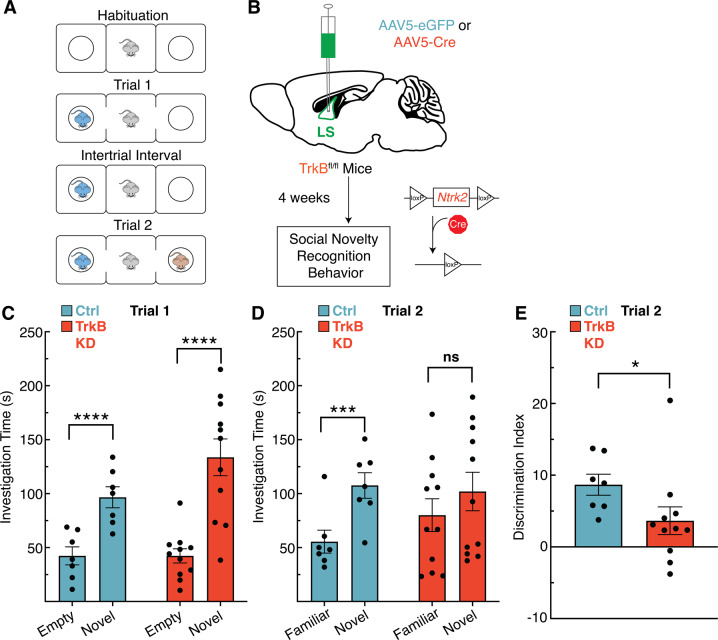
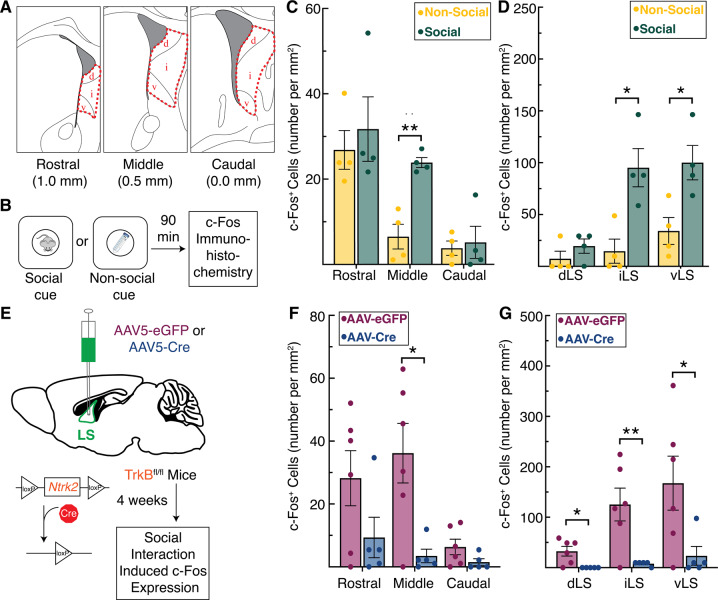
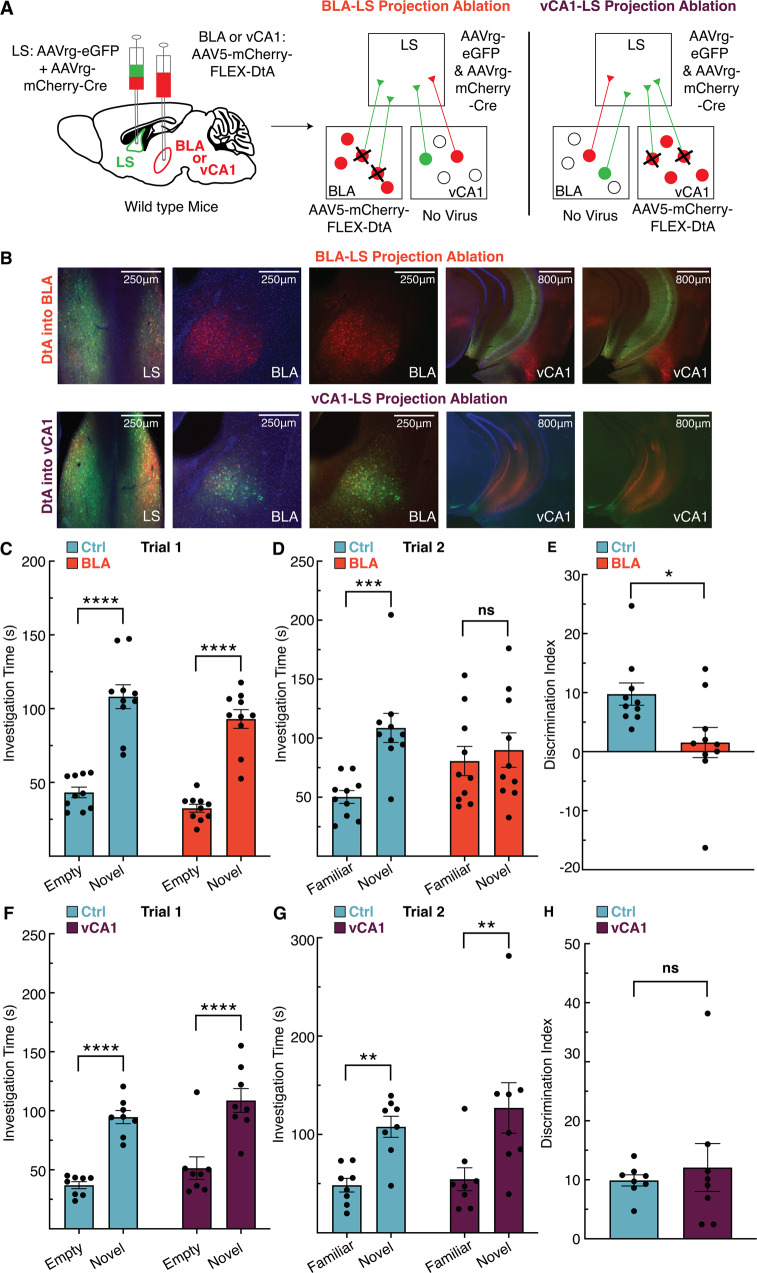
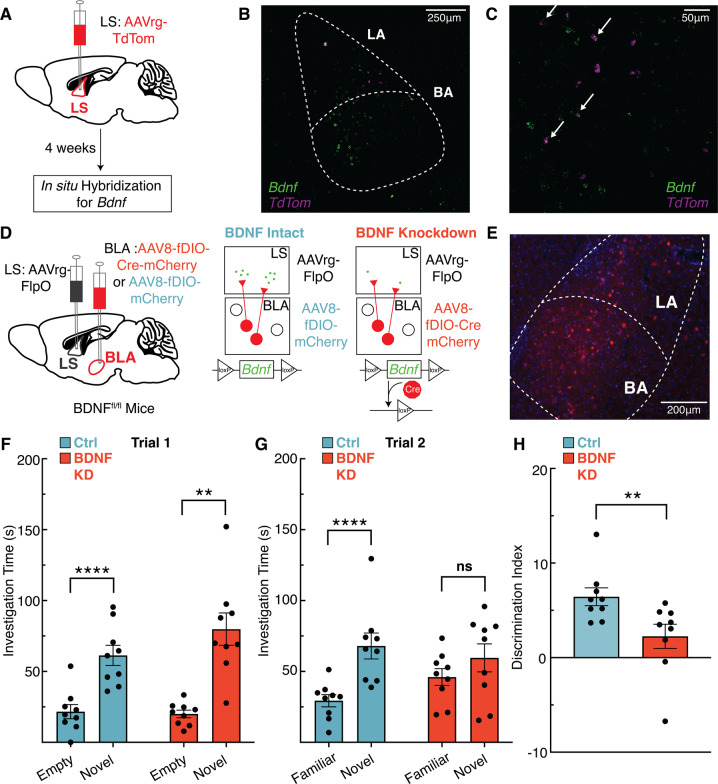
The lateral septum (LS) is a basal forebrain GABAergic region that is implicated in social novelty. However, the neural circuits and cell signaling pathways that converge on the LS to mediate social behaviors aren't well understood. Multiple lines of evidence suggest that signaling of brain-derived neurotrophic factor (BDNF) through its receptor TrkB plays important roles in social behavior. BDNF is not locally produced in LS, but we demonstrate that nearly all LS GABAergic neurons express TrkB. Local TrkB knock-down in LS neurons decreased social novelty recognition and reduced recruitment of neural activity in LS neurons in response to social novelty. Since BDNF is not synthesized in LS, we investigated which inputs to LS could serve as potential BDNF sources for controlling social novelty recognition. We demonstrate that selectively ablating inputs to LS from the basolateral amygdala (BLA), but not from ventral CA1 (vCA1), impairs social novelty recognition. Moreover, depleting BDNF selectively in BLA-LS projection neurons phenocopied the decrease in social novelty recognition caused by either local LS TrkB knockdown or ablation of BLA-LS inputs. These data support the hypothesis that BLA-LS projection neurons serve as a critical source of BDNF for activating TrkB signaling in LS neurons to control social novelty recognition.
© 2022. The Author(s), under exclusive licence to American College of Neuropsychopharmacology.
Conflict of interest statement
KM is the Social Media Editor for
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
