

Influenza A virus reassortment in mammals gives rise to genetically distinct within-host subpopulations
- PMID: 36369504
- PMCID: PMC9652339
- DOI: 10.1038/s41467-022-34611-z
Influenza A virus reassortment in mammals gives rise to genetically distinct within-host subpopulations
Abstract
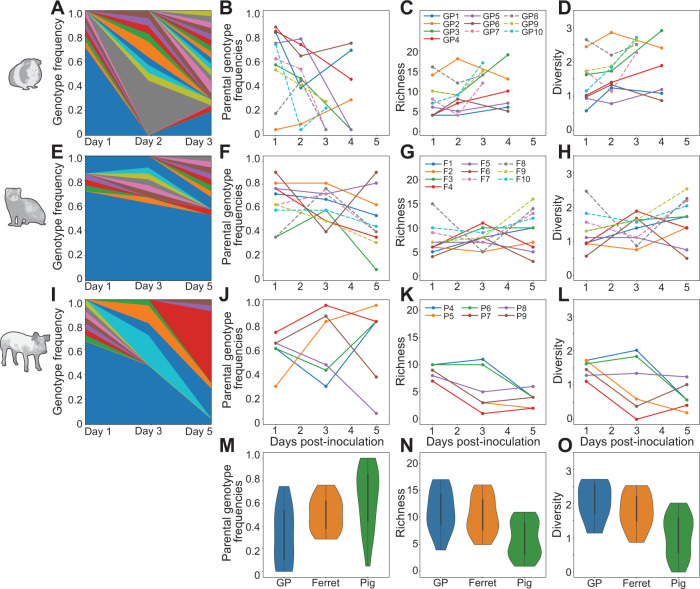
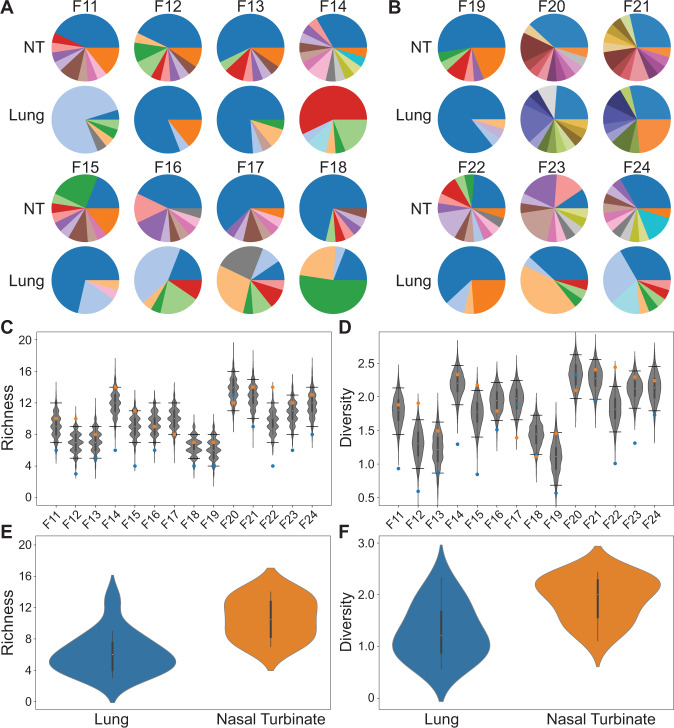
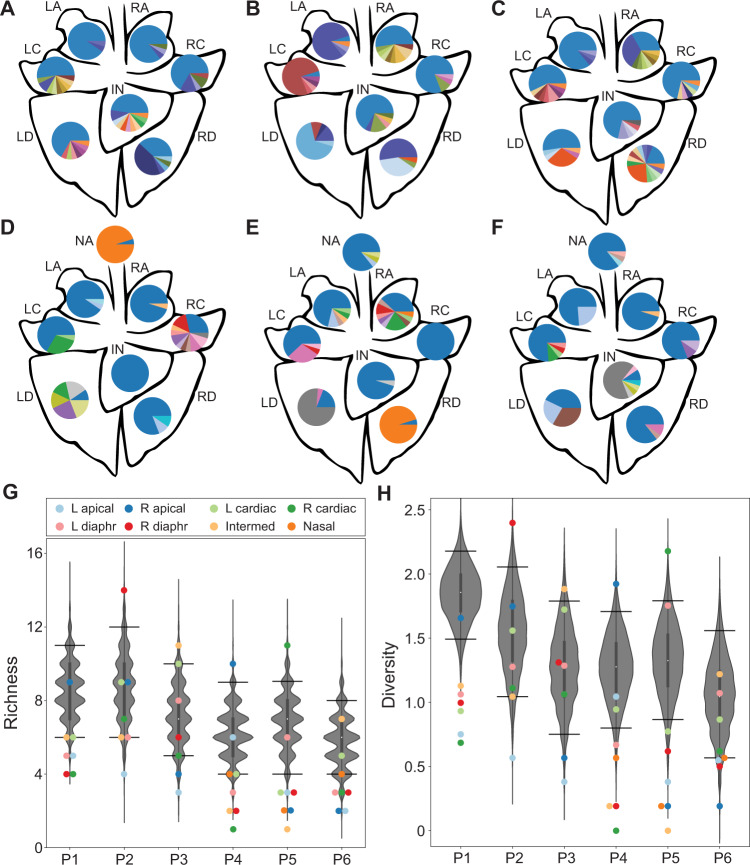
Influenza A virus (IAV) genetic exchange through reassortment has the potential to accelerate viral evolution and has played a critical role in the generation of multiple pandemic strains. For reassortment to occur, distinct viruses must co-infect the same cell. The spatio-temporal dynamics of viral dissemination within an infected host therefore define opportunity for reassortment. Here, we used wild type and synonymously barcoded variant viruses of a pandemic H1N1 strain to examine the within-host viral dynamics that govern reassortment in guinea pigs, ferrets and swine. The first two species are well-established models of human influenza, while swine are a natural host and a frequent conduit for cross-species transmission and reassortment. Our results show reassortment to be pervasive in all three hosts but less frequent in swine than in ferrets and guinea pigs. In ferrets, tissue-specific differences in the opportunity for reassortment are also evident, with more reassortants detected in the nasal tract than the lower respiratory tract. While temporal trends in viral diversity are limited, spatial patterns are clear, with heterogeneity in the viral genotypes detected at distinct anatomical sites revealing extensive compartmentalization of reassortment and replication. Our data indicate that the dynamics of viral replication in mammals allow diversification through reassortment but that the spatial compartmentalization of variants likely shapes their evolution and onward transmission.
© 2022. The Author(s).
Conflict of interest statement
The JAR laboratory received support from Tonix Pharmaceuticals, Xing Technologies, and Zoetis, outside of the reported work. J.A.R. is inventor on patents and patent applications on the use of antivirals and vaccines for the treatment and prevention of virus infections, owned by Kansas State University, KS. The remaining authors declare no competing interests.
Figures
Similar articles
-
Naturally occurring influenza reassortment in pigs facilitates the emergence of intrahost virus subpopulations with distinct genotypes and replicative fitness.mBio. 2025 Jan 8;16(1):e0192424. doi: 10.1128/mbio.01924-24. Epub 2024 Nov 29. mBio. 2025. PMID: 39611844 Free PMC article.
-
Influenza A Virus Reassortment Is Limited by Anatomical Compartmentalization following Coinfection via Distinct Routes.J Virol. 2018 Feb 12;92(5):e02063-17. doi: 10.1128/JVI.02063-17. Print 2018 Mar 1. J Virol. 2018. PMID: 29212934 Free PMC article.
-
Vaccination decreases the risk of influenza A virus reassortment but not genetic variation in pigs.Elife. 2022 Sep 2;11:e78618. doi: 10.7554/eLife.78618. Elife. 2022. PMID: 36052992 Free PMC article.
-
Enhancement of influenza virus transmission by gene reassortment.Curr Top Microbiol Immunol. 2014;385:185-204. doi: 10.1007/82_2014_389. Curr Top Microbiol Immunol. 2014. PMID: 25048543 Review.
-
[Swine influenza virus: evolution mechanism and epidemic characterization--a review].Wei Sheng Wu Xue Bao. 2009 Sep;49(9):1138-45. Wei Sheng Wu Xue Bao. 2009. PMID: 20030049 Review. Chinese.
Cited by
-
Genetic drift and purifying selection shape within-host influenza A virus populations during natural swine infections.PLoS Pathog. 2024 Apr 16;20(4):e1012131. doi: 10.1371/journal.ppat.1012131. eCollection 2024 Apr. PLoS Pathog. 2024. PMID: 38626244 Free PMC article.
-
High-throughput droplet-based analysis of influenza A virus genetic reassortment by single-virus RNA sequencing.Proc Natl Acad Sci U S A. 2023 Feb 7;120(6):e2211098120. doi: 10.1073/pnas.2211098120. Epub 2023 Feb 2. Proc Natl Acad Sci U S A. 2023. PMID: 36730204 Free PMC article.
-
Machine learning methods for predicting human-adaptive influenza A virus reassortment based on intersegment constraint.Front Microbiol. 2025 Mar 21;16:1546536. doi: 10.3389/fmicb.2025.1546536. eCollection 2025. Front Microbiol. 2025. PMID: 40190733 Free PMC article.
-
Zoonotic Animal Influenza Virus and Potential Mixing Vessel Hosts.Viruses. 2023 Apr 16;15(4):980. doi: 10.3390/v15040980. Viruses. 2023. PMID: 37112960 Free PMC article. Review.
-
Respiratory viruses: New frontiers-a Keystone Symposia report.Ann N Y Acad Sci. 2023 Apr;1522(1):60-73. doi: 10.1111/nyas.14958. Epub 2023 Feb 1. Ann N Y Acad Sci. 2023. PMID: 36722473 Free PMC article.
References
-
- Webster RG, Hinshaw VS, Bean WJ, Jr, Turner B, Shortridge KF. Influenza viruses from avian and porcine sources and their possible role in the origin of human pandemic strains. Dev. Biol. Stand. 1977;39:461–468. - PubMed
-
- Wright, P. F., Neumann, G. & Kawaoka, Y. In Fields Virology. Vol. 1 (ed D. M. & Howley Knipe, P. M.) 1691–1740 (Lippincott-Raven, 2006).
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
