

An opioid-gated thalamoaccumbal circuit for the suppression of reward seeking in mice
- PMID: 36369508
- PMCID: PMC9652456
- DOI: 10.1038/s41467-022-34517-w
An opioid-gated thalamoaccumbal circuit for the suppression of reward seeking in mice
Erratum in
-
Author Correction: An opioid-gated thalamoaccumbal circuit for the suppression of reward seeking in mice.Nat Commun. 2023 Aug 7;14(1):4733. doi: 10.1038/s41467-023-40431-6. Nat Commun. 2023. PMID: 37550296 Free PMC article. No abstract available.
Abstract
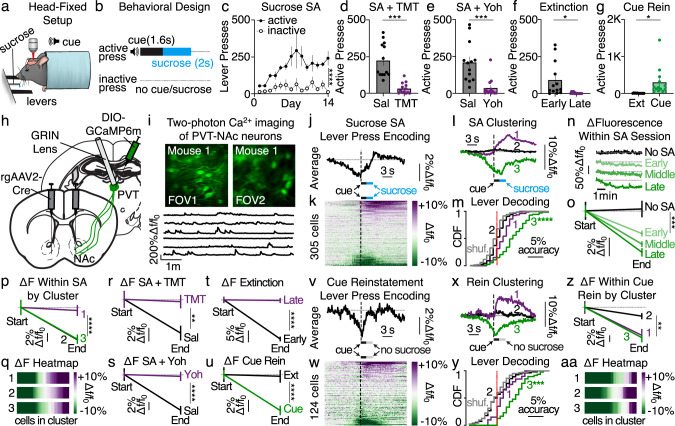
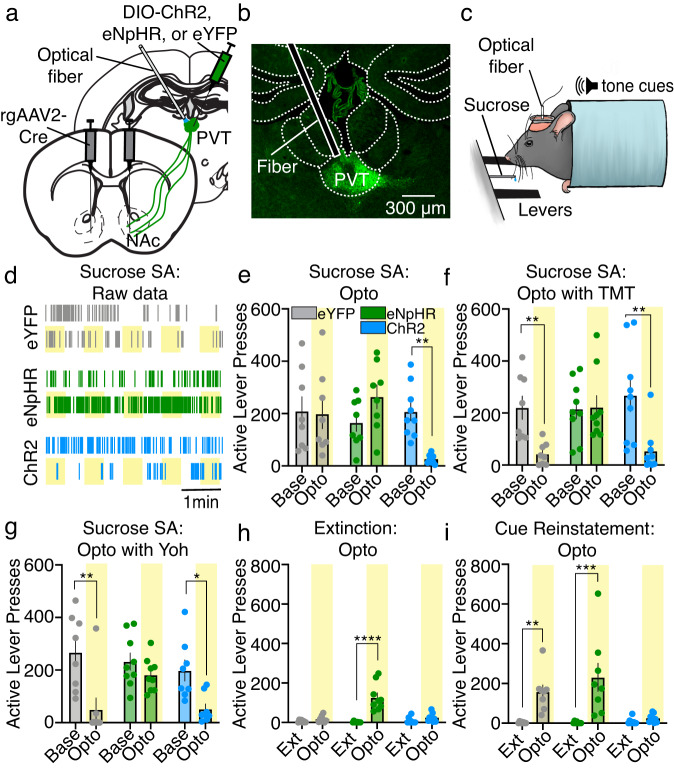
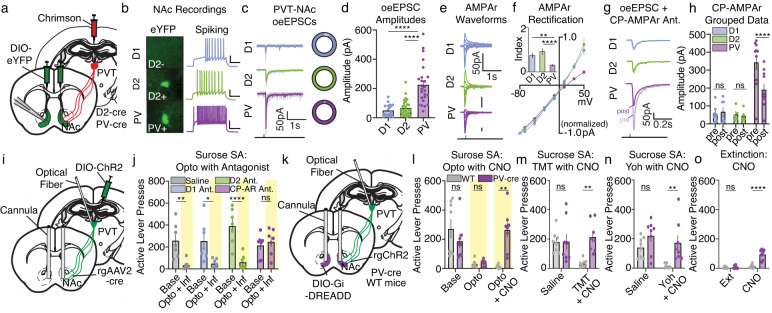
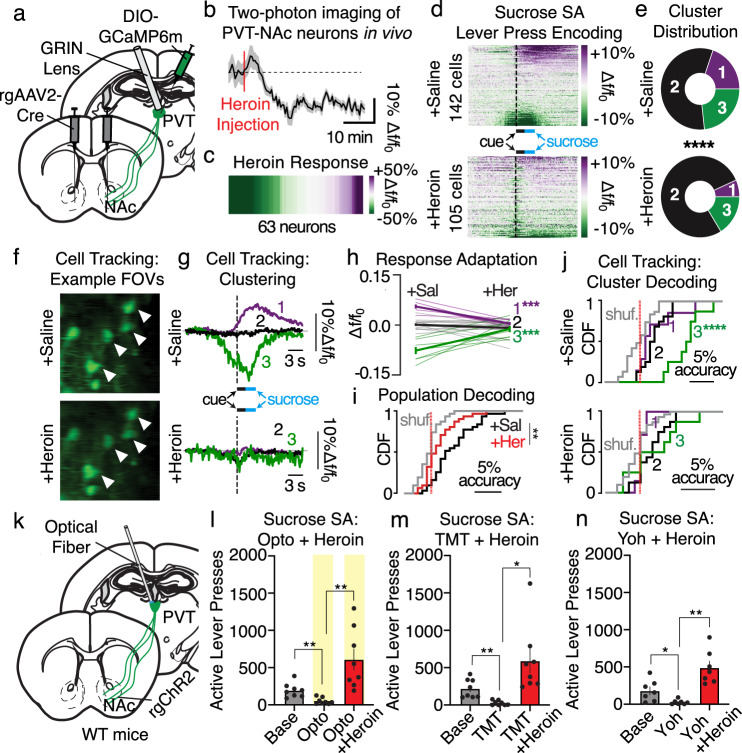
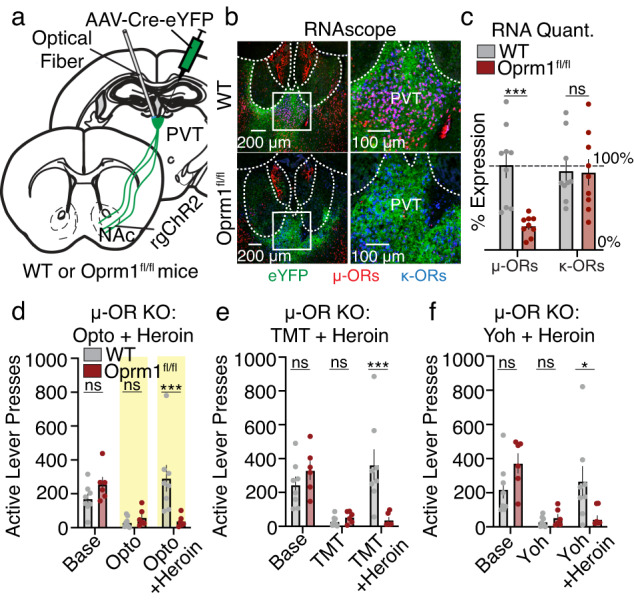
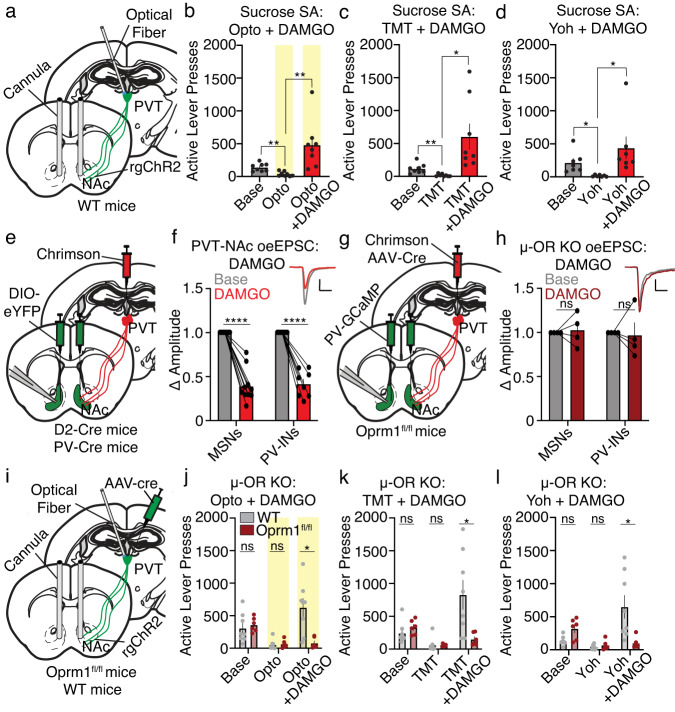
Suppression of dangerous or inappropriate reward-motivated behaviors is critical for survival, whereas therapeutic or recreational opioid use can unleash detrimental behavioral actions and addiction. Nevertheless, the neuronal systems that suppress maladaptive motivated behaviors remain unclear, and whether opioids disengage those systems is unknown. In a mouse model using two-photon calcium imaging in vivo, we identify paraventricular thalamostriatal neuronal ensembles that are inhibited upon sucrose self-administration and seeking, yet these neurons are tonically active when behavior is suppressed by a fear-provoking predator odor, a pharmacological stressor, or inhibitory learning. Electrophysiological, optogenetic, and chemogenetic experiments reveal that thalamostriatal neurons innervate accumbal parvalbumin interneurons through synapses enriched with calcium permeable AMPA receptors, and activity within this circuit is necessary and sufficient for the suppression of sucrose seeking regardless of the behavioral suppressor administered. Furthermore, systemic or intra-accumbal opioid injections rapidly dysregulate thalamostriatal ensemble dynamics, weaken thalamostriatal synaptic innervation of downstream neurons, and unleash reward-seeking behaviors in a manner that is reversed by genetic deletion of thalamic µ-opioid receptors. Overall, our findings reveal a thalamostriatal to parvalbumin interneuron circuit that is both required for the suppression of reward seeking and rapidly disengaged by opioids.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Logan GD. Executive control of thought and action. Acta Psychol. (Amst.) 1985;60:193–210. doi: 10.1016/0001-6918(85)90055-1. - DOI
-
- Stephens, D. W. & Krebs, J. R. Foraging Theory. (Princeton University Press, 1986).
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
