

Loss of hypothalamic Furin affects POMC to proACTH cleavage and feeding behavior in high-fat diet-fed mice
- PMID: 36374777
- PMCID: PMC9664468
- DOI: 10.1016/j.molmet.2022.101627
Loss of hypothalamic Furin affects POMC to proACTH cleavage and feeding behavior in high-fat diet-fed mice
Abstract
Objective: The hypothalamus regulates feeding and glucose homeostasis through the balanced action of different neuropeptides, which are cleaved and activated by the proprotein convertases PC1/3 and PC2. However, the recent association of polymorphisms in the proprotein convertase FURIN with type 2 diabetes, metabolic syndrome, and obesity, prompted us to investigate the role of FURIN in hypothalamic neurons controlling glucose and feeding.
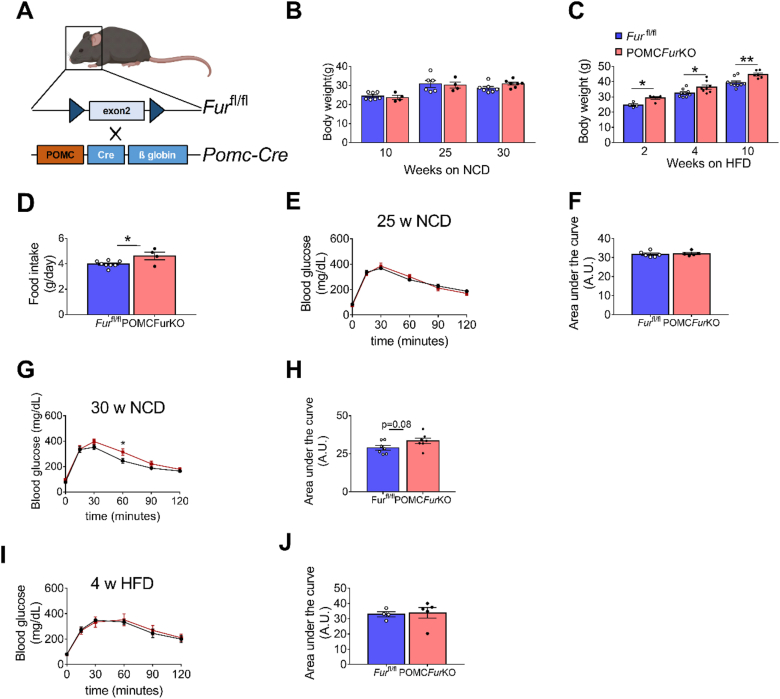
Methods: POMC-Cre+/- mice were bred with Furinfl/fl mice to generate conditional knockout mice with Furin-deletion in neurons expressing proopiomelanocortin (POMCFurKO), and Furinfl/fl mice were used as controls. POMCFurKO and controls were periodically monitored on both normal chow diet and high fat diet (HFD) for body weight and glucose tolerance by established in-vivo procedures. Food intake was measured in HFD-fed FurKO and controls. Hypothalamic Pomc mRNA was measured by RT-qPCR. ELISAs quantified POMC protein and resulting peptides in the hypothalamic extracts of POMCFurKO mice and controls. The in-vitro processing of POMC was studied by biochemical techniques in HEK293T and CHO cell lines lacking FURIN.
Results: In control mice, Furin mRNA levels were significantly upregulated on HFD feeding, suggesting an increased demand for FURIN activity in obesogenic conditions. Under these conditions, the POMCFurKO mice were hyperphagic and had increased body weight compared to Furinfl/fl mice. Moreover, protein levels of POMC were elevated and ACTH concentrations markedly reduced. Also, the ratio of α-MSH/POMC was decreased in POMCFurKO mice compared to controls. This indicates that POMC processing was significantly reduced in the hypothalami of POMCFurKO mice, highlighting for the first time the involvement of FURIN in the cleavage of POMC. Importantly, we found that in vitro, the first stage in processing where POMC is cleaved into proACTH was achieved by FURIN but not by PC1/3 or the other proprotein convertases in cell lines lacking a regulated secretory pathway.
Conclusions: These results suggest that FURIN processes POMC into proACTH before sorting into the regulated secretory pathway, challenging the dogma that PC1/3 and PC2 are the only convertases responsible for POMC cleavage. Furthermore, its deletion affects feeding behaviors under obesogenic conditions.
Keywords: ACTH; FURIN; Hypothalamus; Obesity; POMC; proACTH.
Copyright © 2022 The Authors. Published by Elsevier GmbH.. All rights reserved.
Figures



References
-
- Myers M.G., Olson D.P. Central nervous system control of metabolism. Nature. 2012 15;491(7424):357–363. - PubMed
-
- Zhang L., Hernandez-Sanchez D., Herzog H. Regulation of feeding-related behaviors by arcuate neuropeptide Y neurons. Endocrinology. 2019;160(6):1411–1420. - PubMed
-
- Cowley M.A., Smart J.L., Rubinstein M., Cerdán M.G., Diano S., Horvath T.L., et al. Leptin activates anorexigenic POMC neurons through a neural network in the arcuate nucleus. Nature. 2001;411(6836):480–484. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
