

Systematic proximal mapping of the classical RAD51 paralogs unravel functionally and clinically relevant interactors for genome stability
- PMID: 36374936
- PMCID: PMC9718398
- DOI: 10.1371/journal.pgen.1010495
Systematic proximal mapping of the classical RAD51 paralogs unravel functionally and clinically relevant interactors for genome stability
Abstract
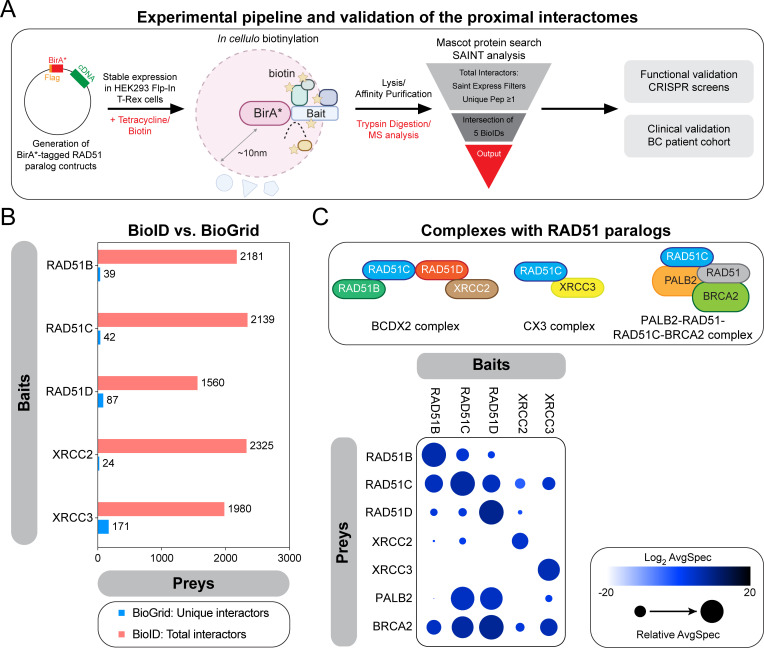
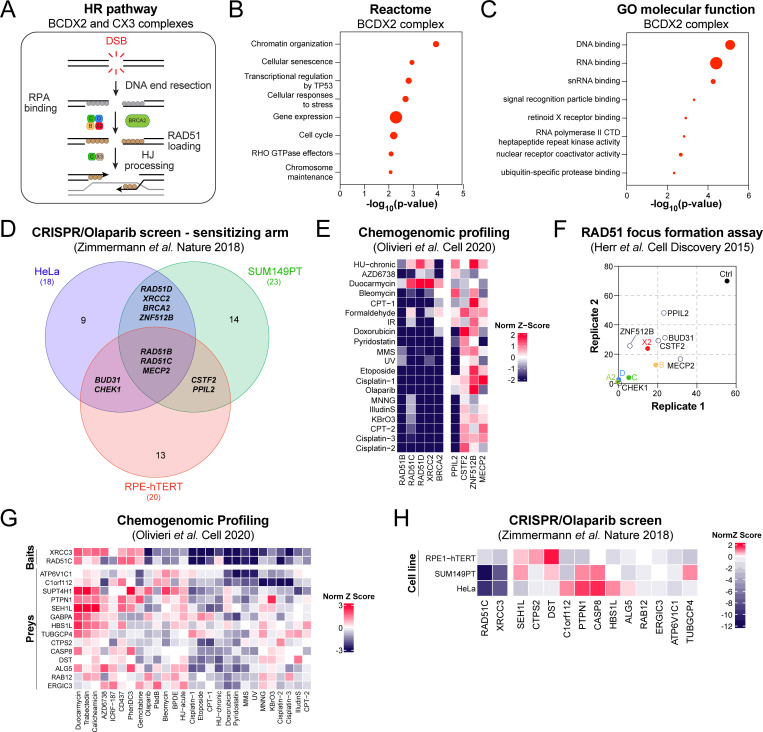
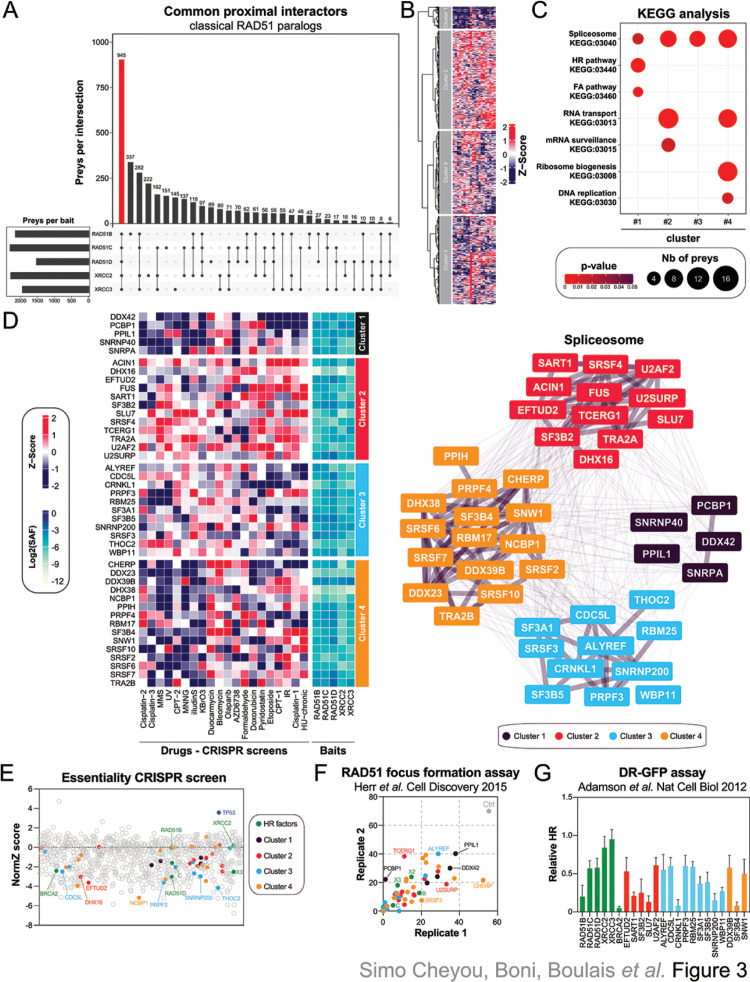
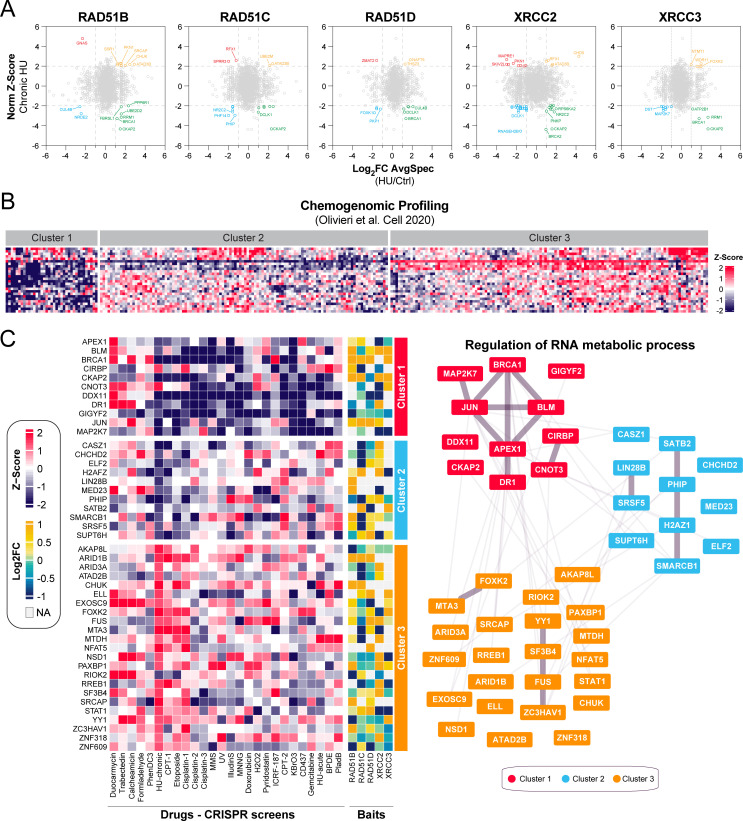
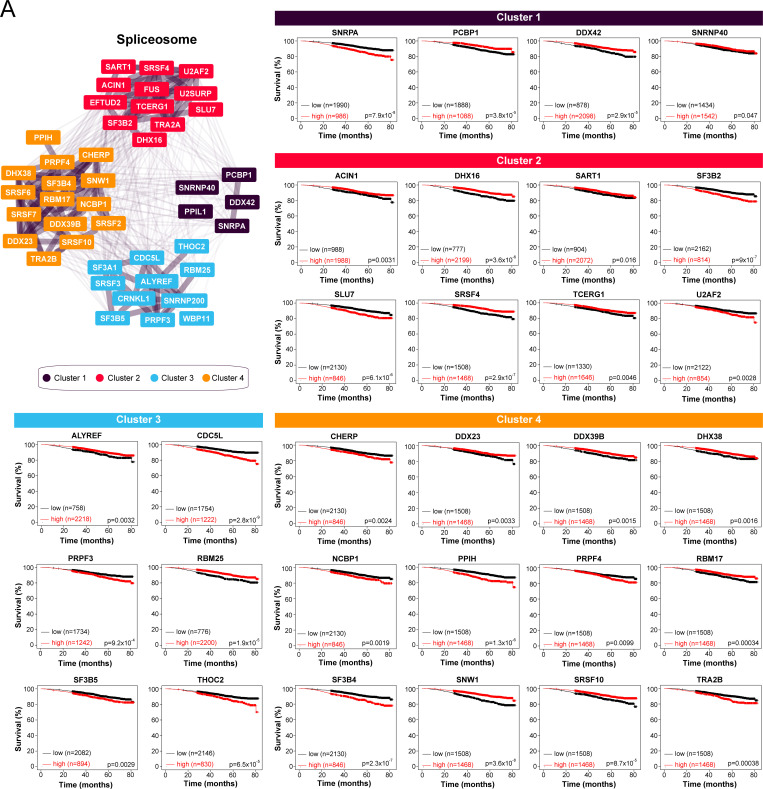
Homologous recombination (HR) plays an essential role in the maintenance of genome stability by promoting the repair of cytotoxic DNA double strand breaks (DSBs). More recently, the HR pathway has emerged as a core component of the response to replication stress, in part by protecting stalled replication forks from nucleolytic degradation. In that regard, the mammalian RAD51 paralogs (RAD51B, RAD51C, RAD51D, XRCC2, and XRCC3) have been involved in both HR-mediated DNA repair and collapsed replication fork resolution. Still, it remains largely obscure how they participate in both processes, thereby maintaining genome stability and preventing cancer development. To gain better insight into their contribution in cellulo, we mapped the proximal interactome of the classical RAD51 paralogs using the BioID approach. Aside from identifying the well-established BCDX2 and CX3 sub-complexes, the spliceosome machinery emerged as an integral component of our proximal mapping, suggesting a crosstalk between this pathway and the RAD51 paralogs. Furthermore, we noticed that factors involved RNA metabolic pathways are significantly modulated within the BioID of the classical RAD51 paralogs upon exposure to hydroxyurea (HU), pointing towards a direct contribution of RNA processing during replication stress. Importantly, several members of these pathways have prognostic potential in breast cancer (BC), where their RNA expression correlates with poorer patient outcome. Collectively, this study uncovers novel functionally relevant partners of the different RAD51 paralogs in the maintenance of genome stability that could be used as biomarkers for the prognosis of BC.
Copyright: © 2022 Simo Cheyou et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
