

Systems-Wide Dissection of Organic Acid Assimilation in Pseudomonas aeruginosa Reveals a Novel Path To Underground Metabolism
- PMID: 36377867
- PMCID: PMC9765439
- DOI: 10.1128/mbio.02541-22
Systems-Wide Dissection of Organic Acid Assimilation in Pseudomonas aeruginosa Reveals a Novel Path To Underground Metabolism
Abstract
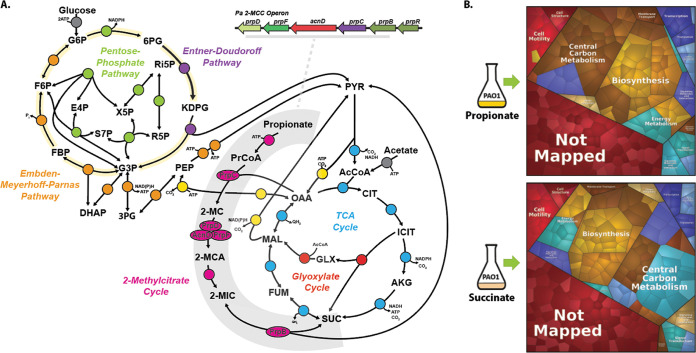
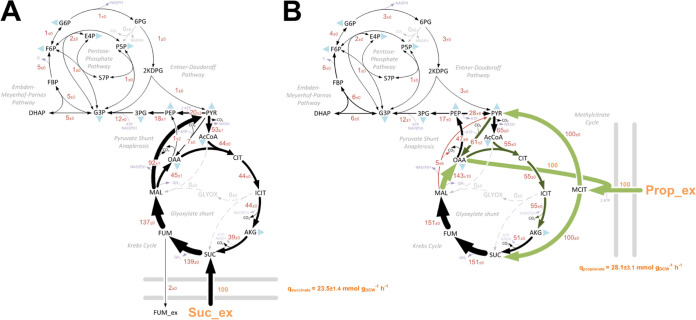
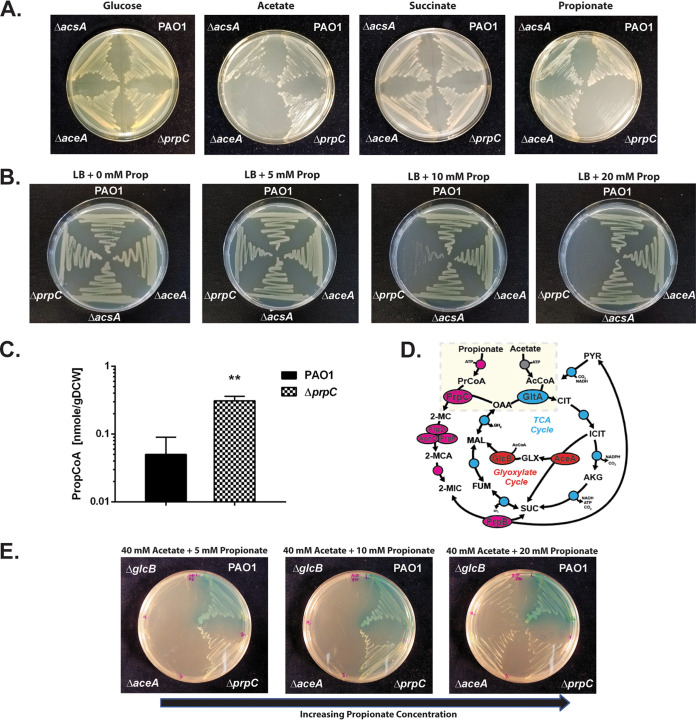
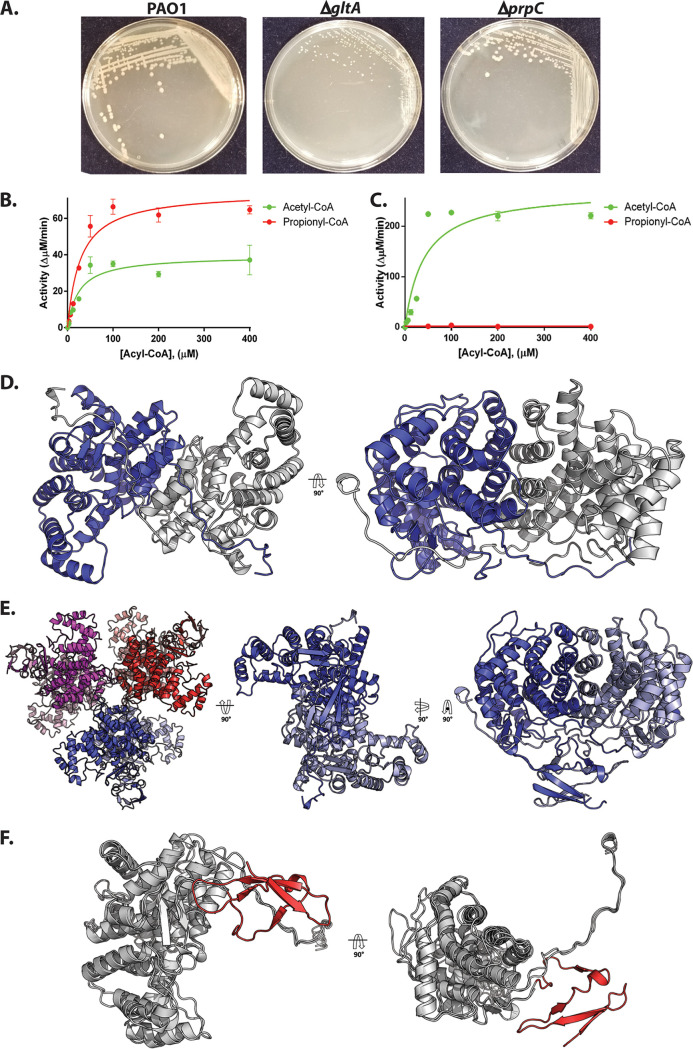
The human pathogen Pseudomonas aeruginosa (Pa) is one of the most frequent and severe causes of nosocomial infection. This organism is also a major cause of airway infections in people with cystic fibrosis (CF). Pa is known to have a remarkable metabolic plasticity, allowing it to thrive under diverse environmental conditions and ecological niches; yet, little is known about the central metabolic pathways that sustain its growth during infection or precisely how these pathways operate. In this work, we used a combination of 'omics approaches (transcriptomics, proteomics, metabolomics, and 13C-fluxomics) and reverse genetics to provide systems-level insight into how the infection-relevant organic acids succinate and propionate are metabolized by Pa. Moreover, through structural and kinetic analysis of the 2-methylcitrate synthase (2-MCS; PrpC) and its paralogue citrate (CIT) synthase (GltA), we show how these two crucial enzymatic steps are interconnected in Pa organic acid assimilation. We found that Pa can rapidly adapt to the loss of GltA function by acquiring mutations in a transcriptional repressor, which then derepresses prpC expression. Our findings provide a clear example of how "underground metabolism," facilitated by enzyme substrate promiscuity, "rewires" Pa metabolism, allowing it to overcome the loss of a crucial enzyme. This pathogen-specific knowledge is critical for the advancement of a model-driven framework to target bacterial central metabolism. IMPORTANCE Pseudomonas aeruginosa is an opportunistic human pathogen that, due to its unrivalled resistance to antibiotics, ubiquity in the built environment, and aggressiveness in infection scenarios, has acquired the somewhat dubious accolade of being designated a "critical priority pathogen" by the WHO. In this work, we uncover the pathways and mechanisms used by P. aeruginosa to grow on a substrate that is abundant at many infection sites: propionate. We found that if the organism is prevented from metabolizing propionate, the substrate turns from being a convenient nutrient source into a potent poison, preventing bacterial growth. We further show that one of the enzymes involved in these reactions, 2-methylcitrate synthase (PrpC), is promiscuous and can moonlight for another essential enzyme in the cell (citrate synthase). Indeed, mutations that abolish citrate synthase activity (which would normally prevent the cell from growing) can be readily overcome if the cell acquires additional mutations that increase the expression of PrpC. This is a nice example of the evolutionary utility of so-called "underground metabolism."
Keywords: 2-methylcitrate cycle; Pseudomonas aeruginosa; central metabolism; enzyme promiscuity; propionate metabolism; underground metabolism.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
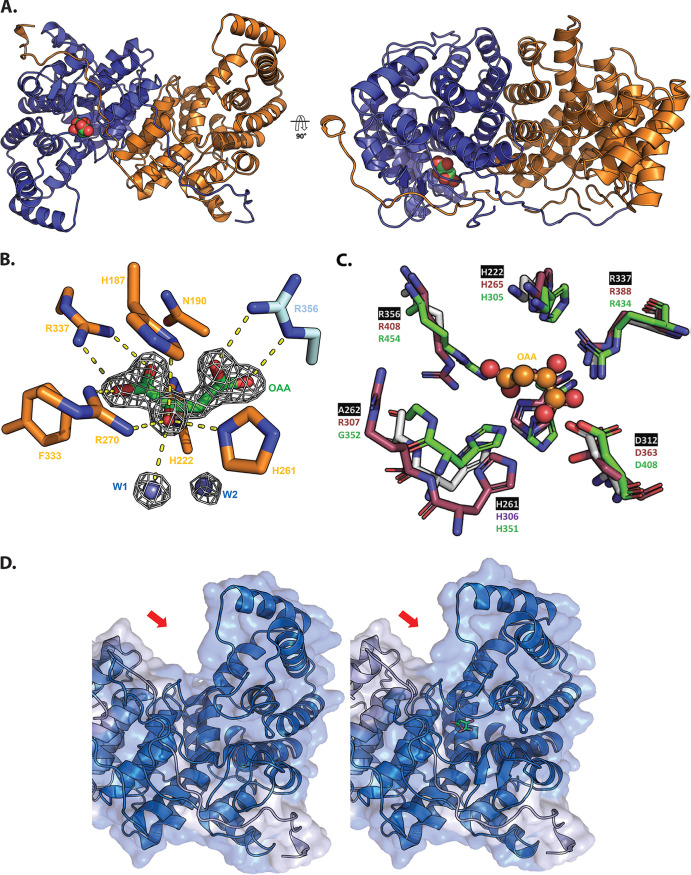
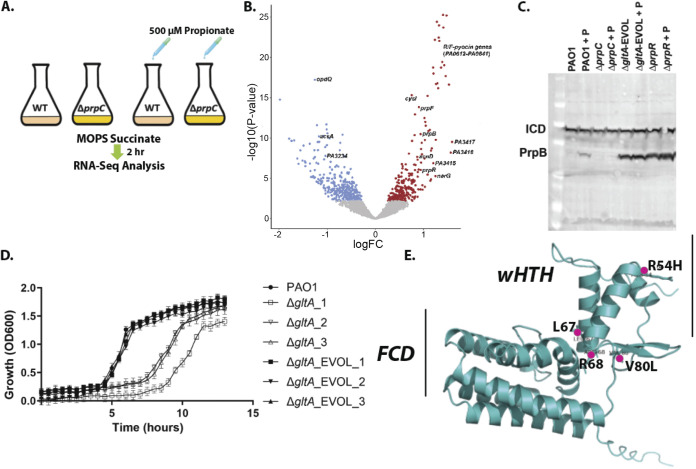
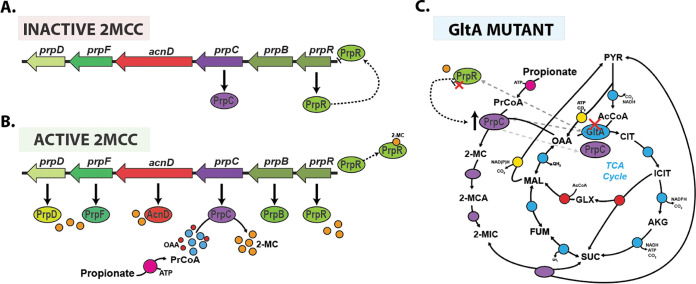
References
-
- Sun Z, Kang Y, Norris MH, Troyer RM, Son MS, Schweizer HP, Dow SW, Hoang TT. 2014. Blocking phosphatidylcholine utilization in Pseudomonas aeruginosa, via mutagenesis of fatty acid, glycerol and choline degradation pathways, confirms the importance of this nutrient source in vivo. PLoS One 9:e103778. doi: 10.1371/journal.pone.0103778. - DOI - PMC - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
