

Aging drives Tet2+/- clonal hematopoiesis via IL-1 signaling
- PMID: 36379023
- PMCID: PMC10651783
- DOI: 10.1182/blood.2022016835
Aging drives Tet2+/- clonal hematopoiesis via IL-1 signaling
Abstract
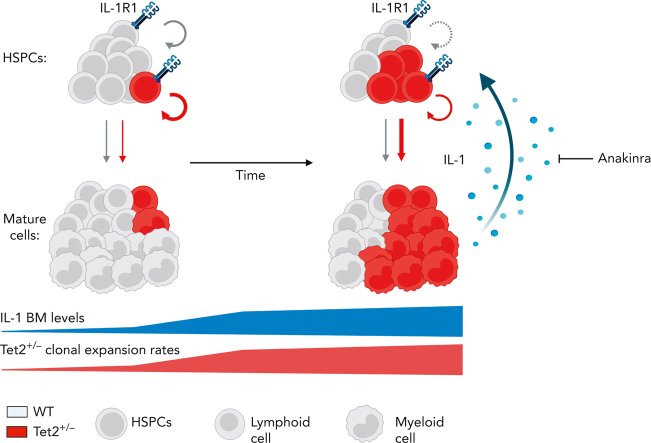
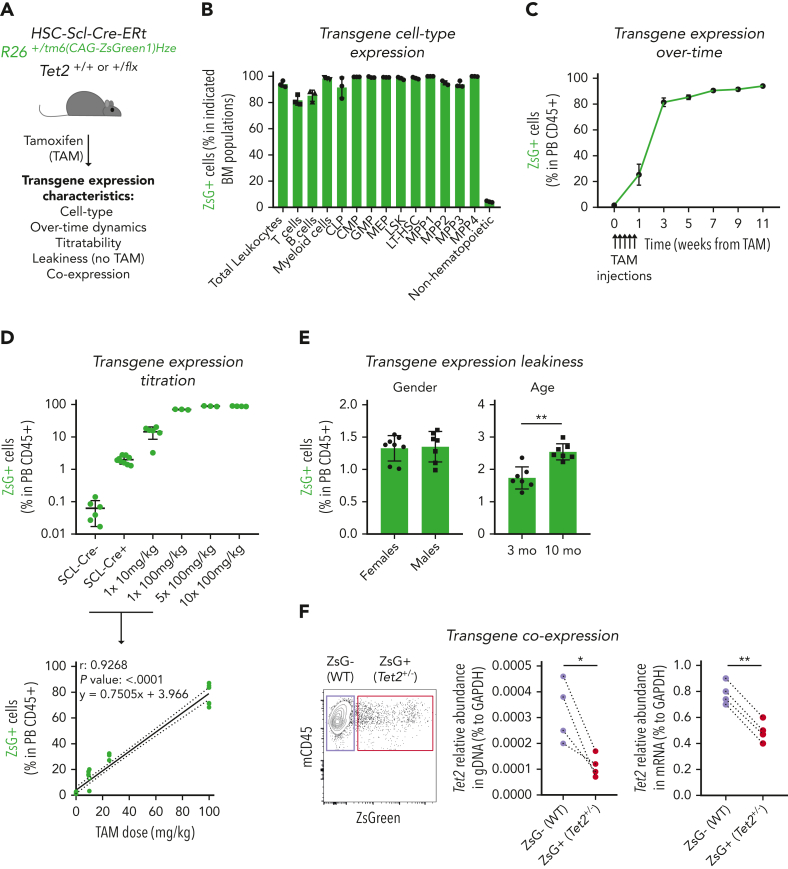
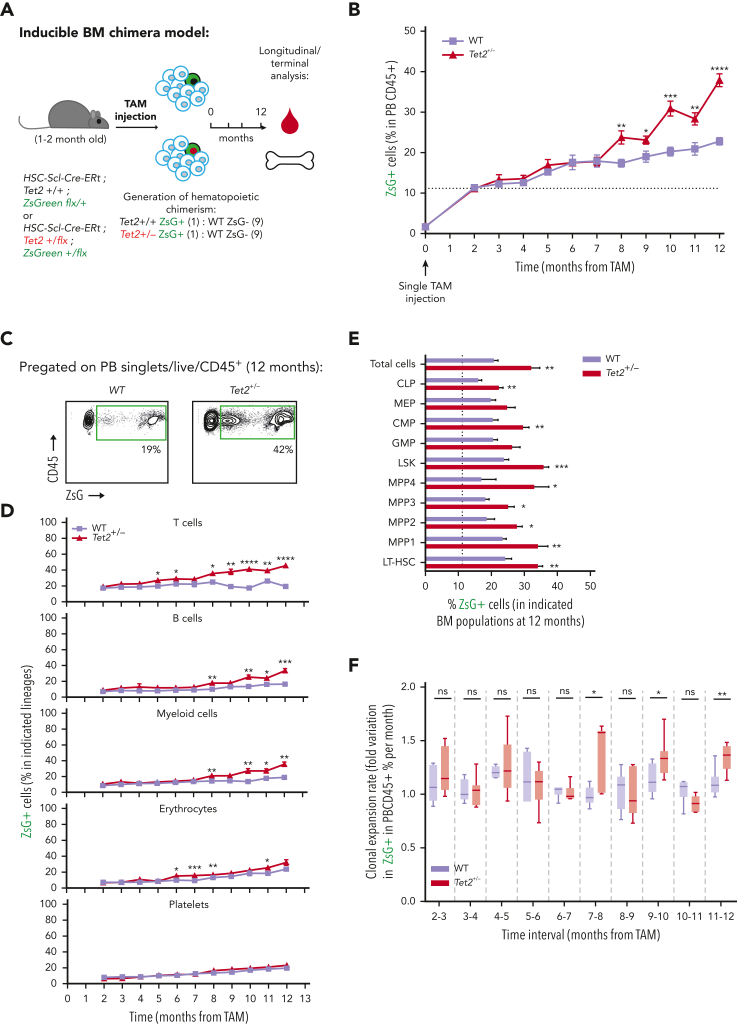
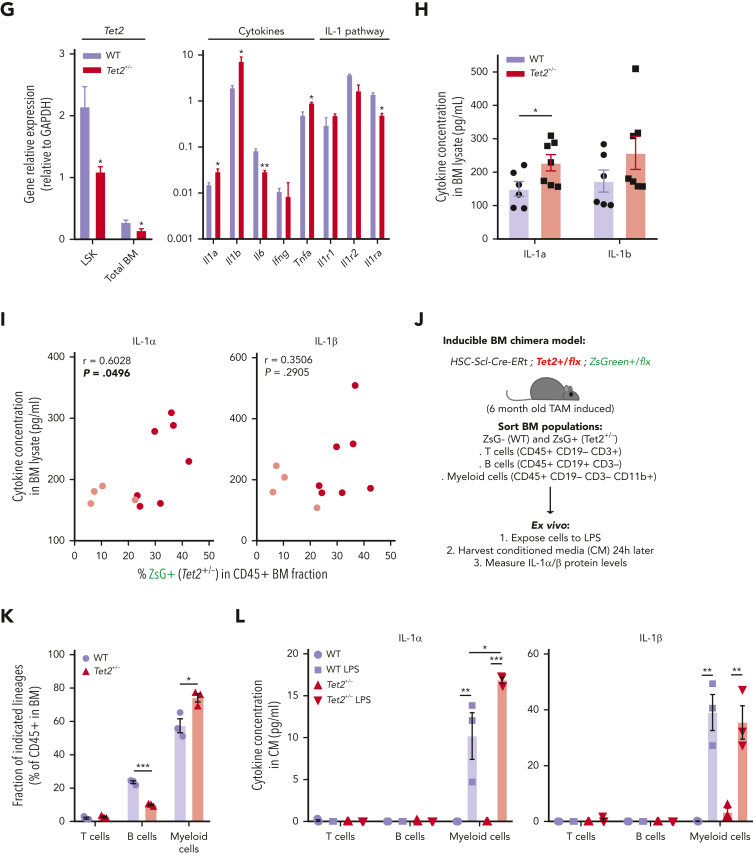
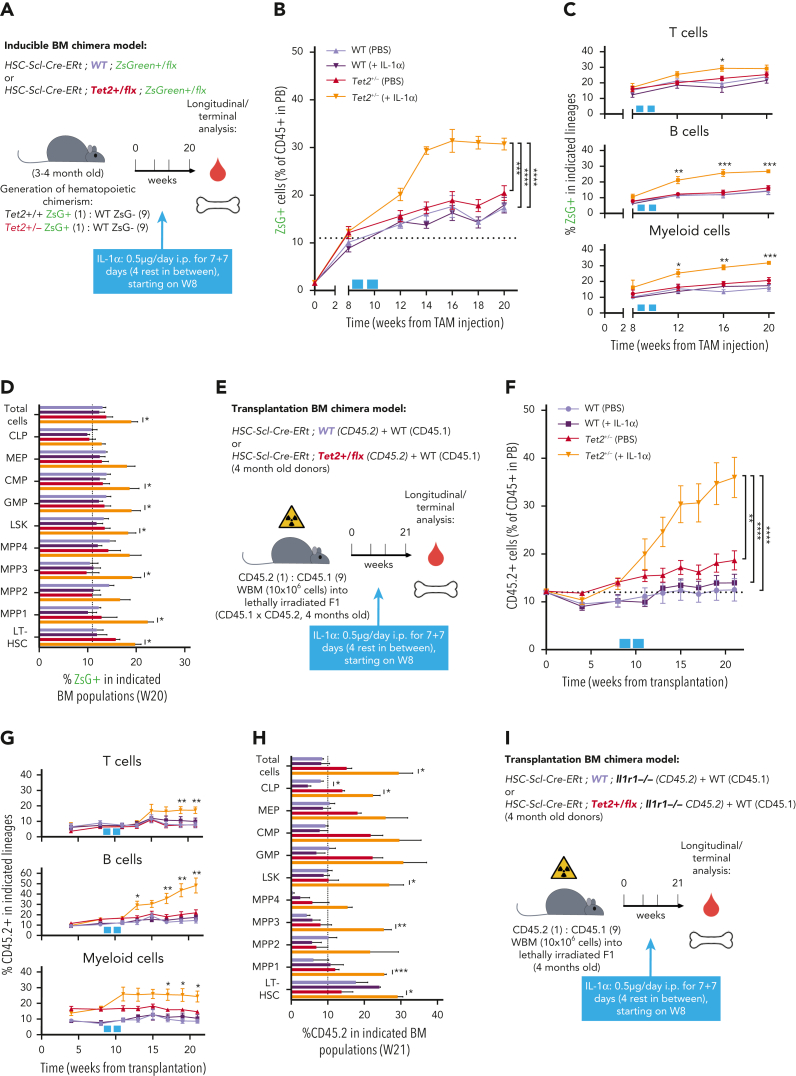
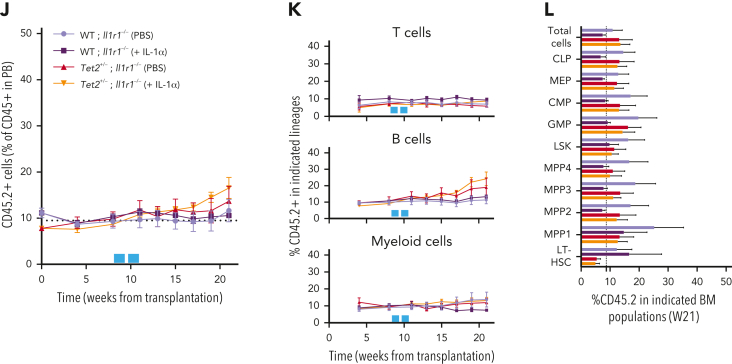
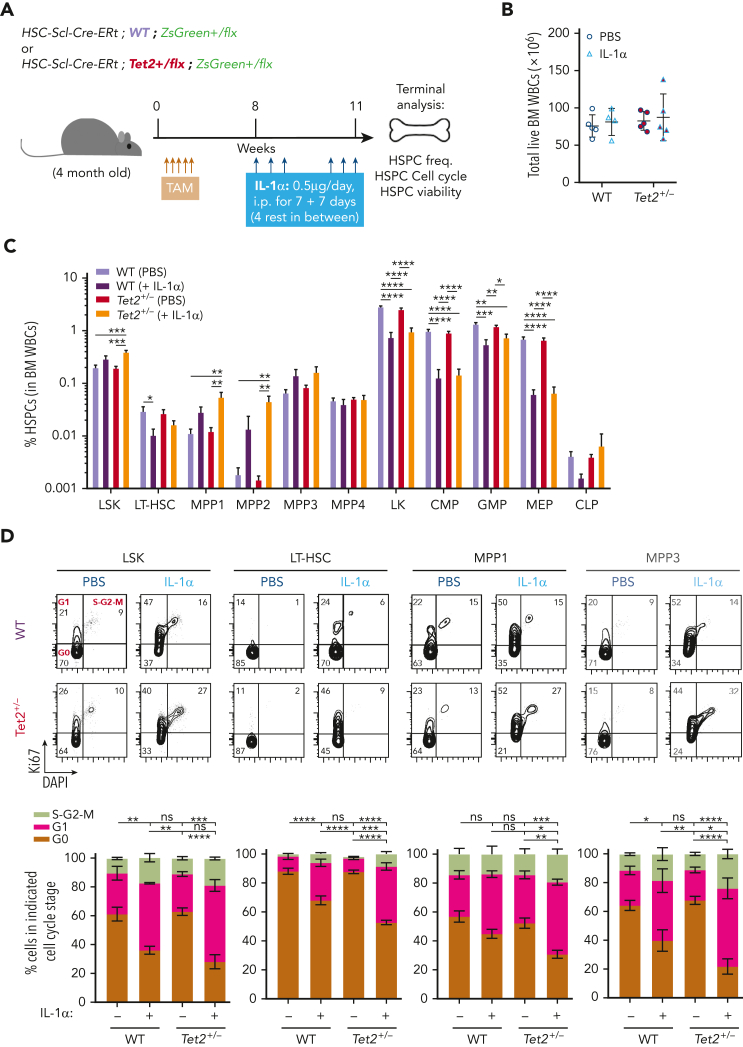
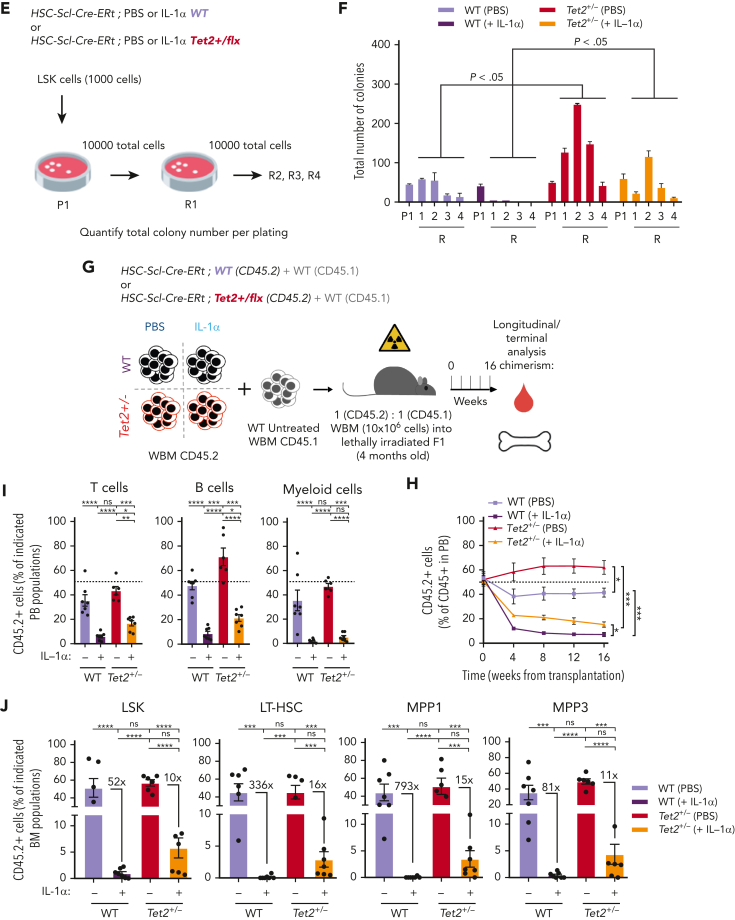
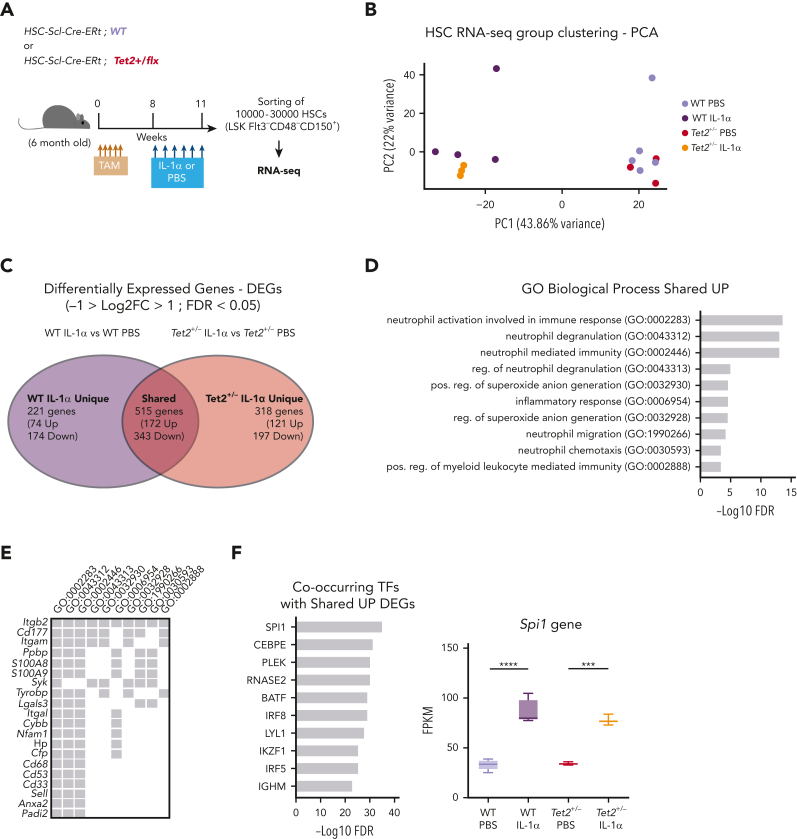
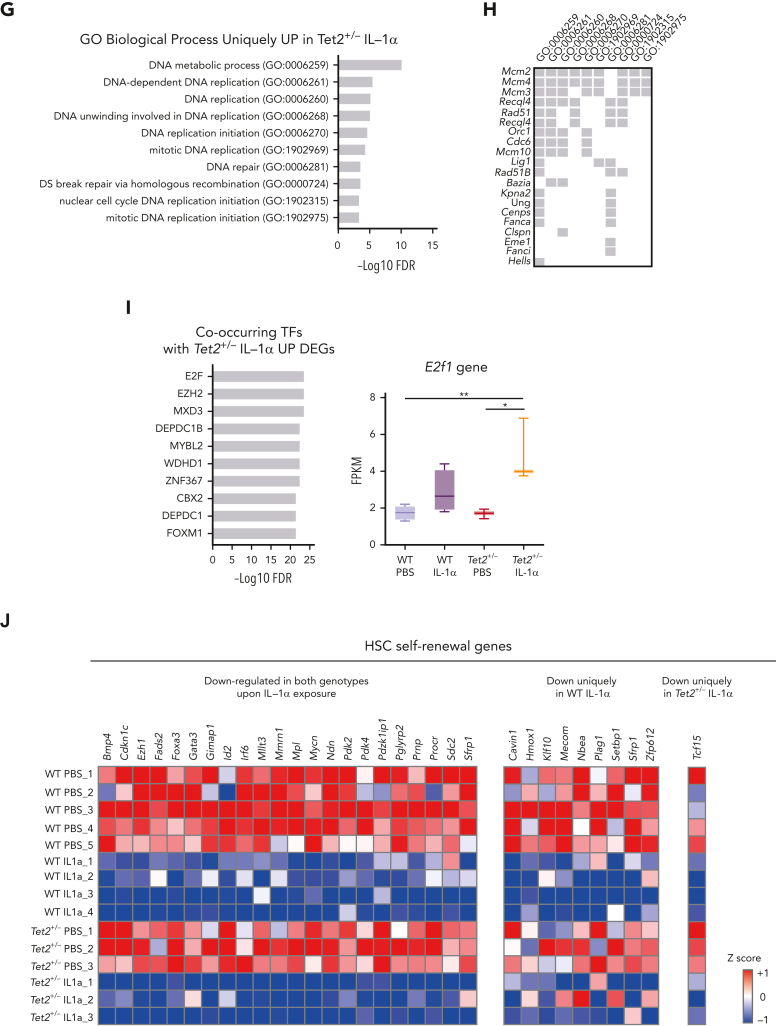
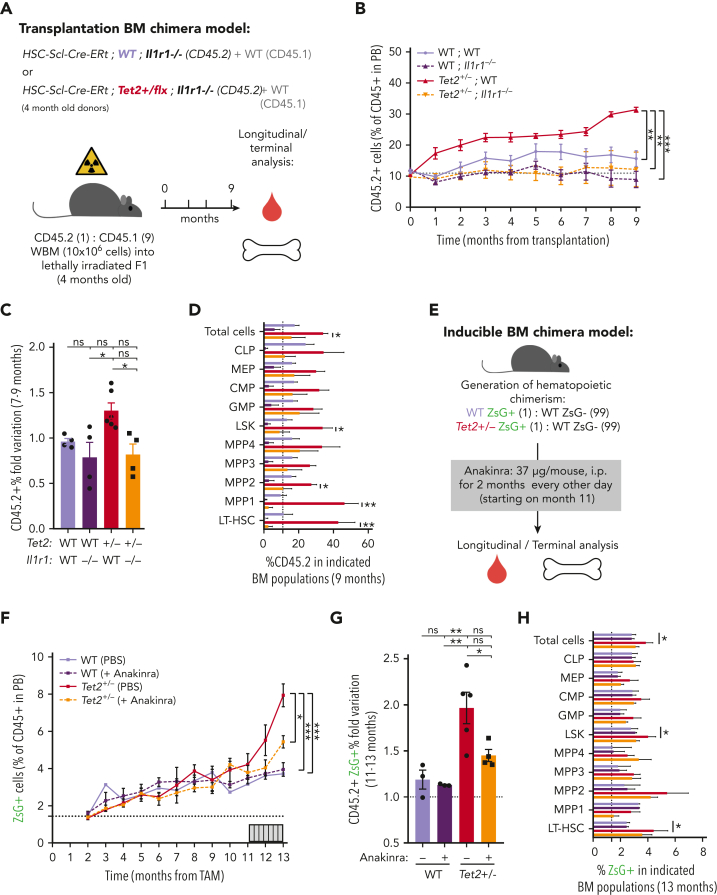
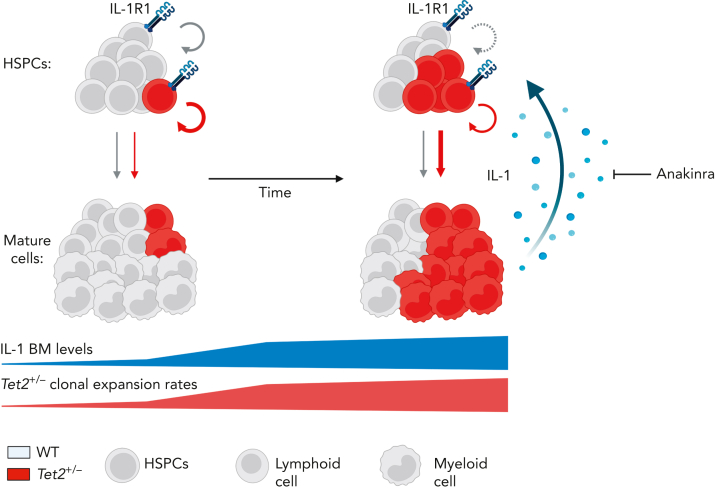
Clonal hematopoiesis of indeterminate potential (CHIP), also referred to as aging-related clonal hematopoiesis, is defined as an asymptomatic clonal expansion of mutant mature hematopoietic cells in ≥4% of blood leukocytes. CHIP associates with advanced age and increased risk for hematological malignancy, cardiovascular disease, and all-cause mortality. Loss-of-function somatic mutations in TET2 are frequent drivers of CHIP. However, the contribution of aging-associated cooperating cell-extrinsic drivers, like inflammation, remains underexplored. Using bone marrow (BM) transplantation and newly developed genetic mosaicism (HSC-SCL-Cre-ERT; Tet2+/flox; R26+/tm6[CAG-ZsGreen1]Hze) mouse models of Tet2+/-driven CHIP, we observed an association between increased Tet2+/- clonal expansion and higher BM levels of the inflammatory cytokine interleukin-1 (IL-1) upon aging. Administration of IL-1 to mice carrying CHIP led to an IL-1 receptor 1 (IL-1R1)-dependent expansion of Tet2+/- hematopoietic stem and progenitor cells (HSPCs) and mature blood cells. This expansion was caused by increased Tet2+/- HSPC cell cycle progression, increased multilineage differentiation, and higher repopulation capacity compared with their wild-type counterparts. In agreement, IL-1α-treated Tet2+/- hematopoietic stem cells showed increased DNA replication and repair transcriptomic signatures and reduced susceptibility to IL-1α-mediated downregulation of self-renewal genes. More important, genetic deletion of IL-1R1 in Tet2+/- HPSCs or pharmacologic inhibition of IL-1 signaling impaired Tet2+/- clonal expansion, establishing the IL-1 pathway as a relevant and therapeutically targetable driver of Tet2+/- CHIP progression during aging.
© 2023 by The American Society of Hematology. Licensed under Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International (CC BY-NC-ND 4.0), permitting only noncommercial, nonderivative use with attribution. All other rights reserved.
Conflict of interest statement
Conflict-of-interest disclosure: The authors declare no competing financial interests.
Figures
Comment in
-
Breaking the CH inflammation-expansion cycle.Blood. 2023 Feb 23;141(8):815-816. doi: 10.1182/blood.2022019104. Blood. 2023. PMID: 36821185 No abstract available.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
