

The miR-124-AMPAR pathway connects polygenic risks with behavioral changes shared between schizophrenia and bipolar disorder
- PMID: 36379214
- PMCID: PMC10183200
- DOI: 10.1016/j.neuron.2022.10.031
The miR-124-AMPAR pathway connects polygenic risks with behavioral changes shared between schizophrenia and bipolar disorder
Abstract
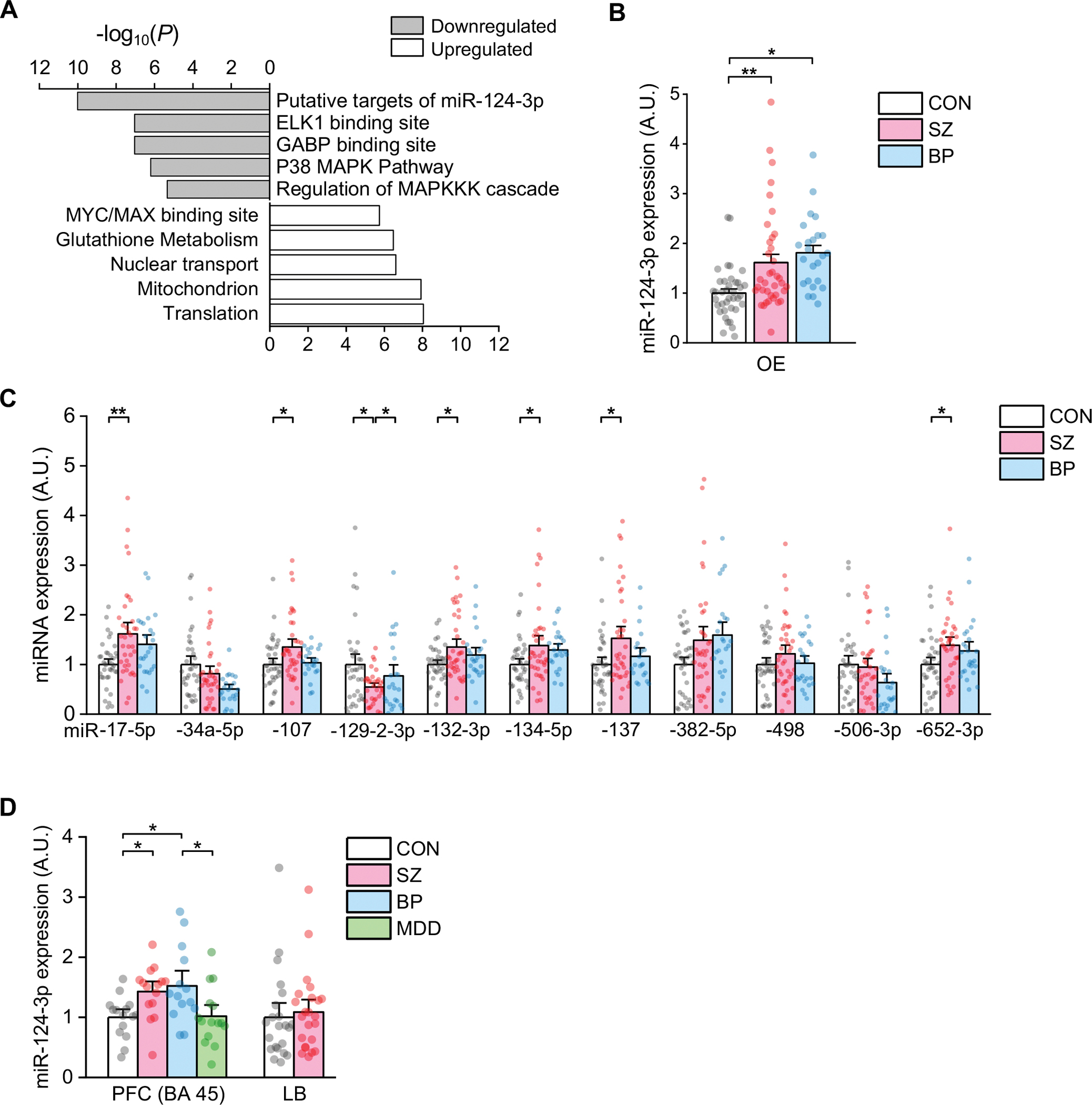
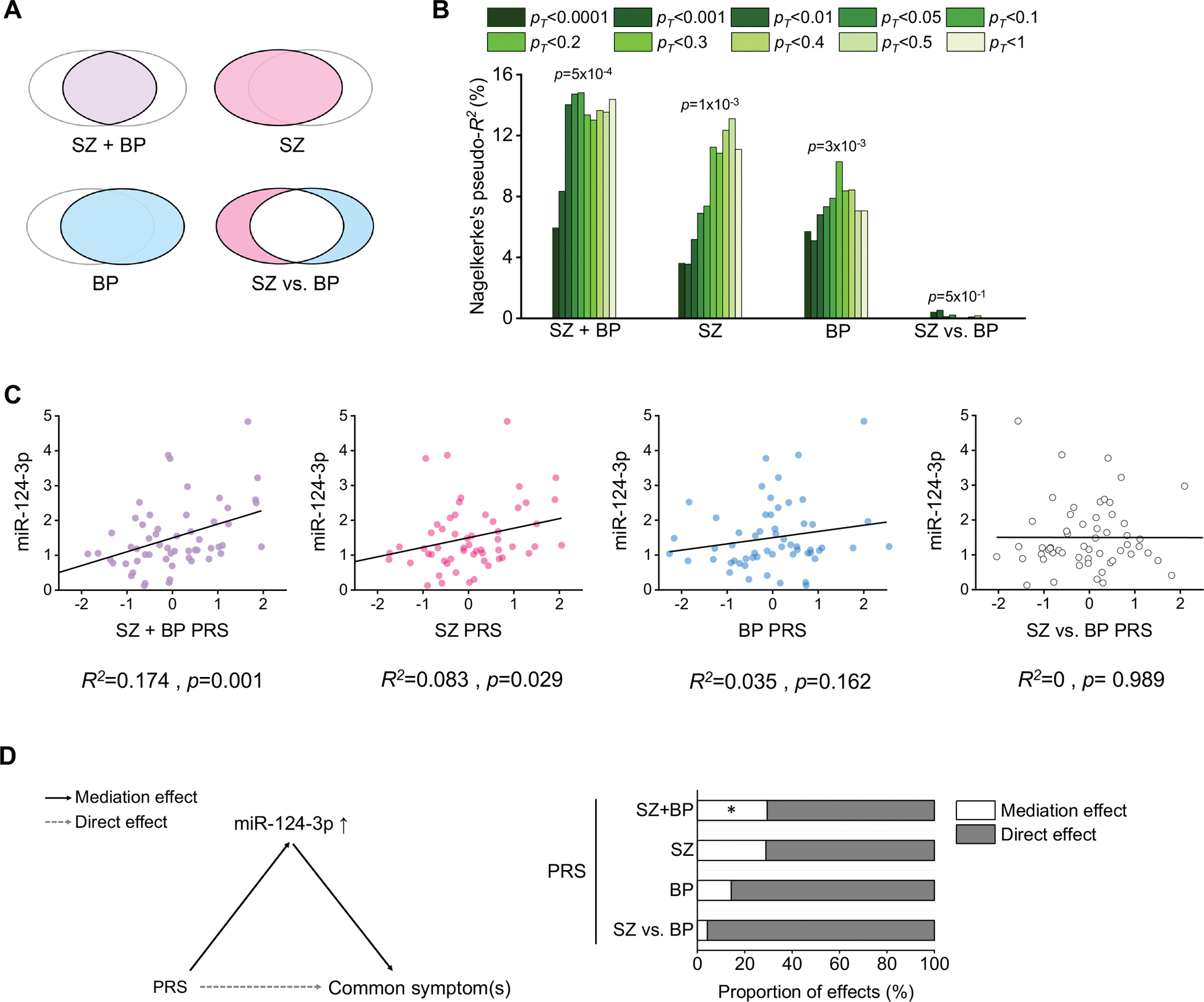
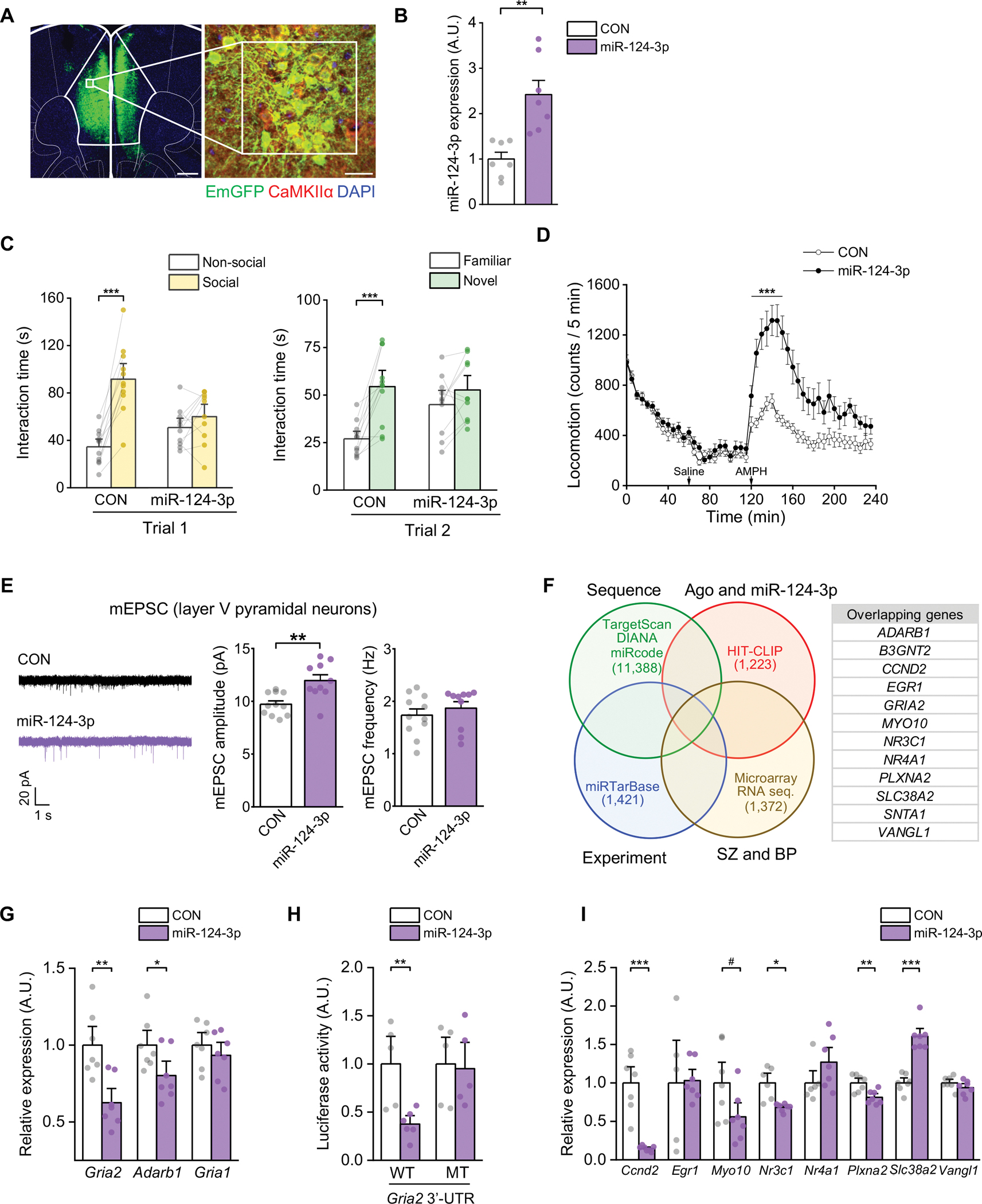
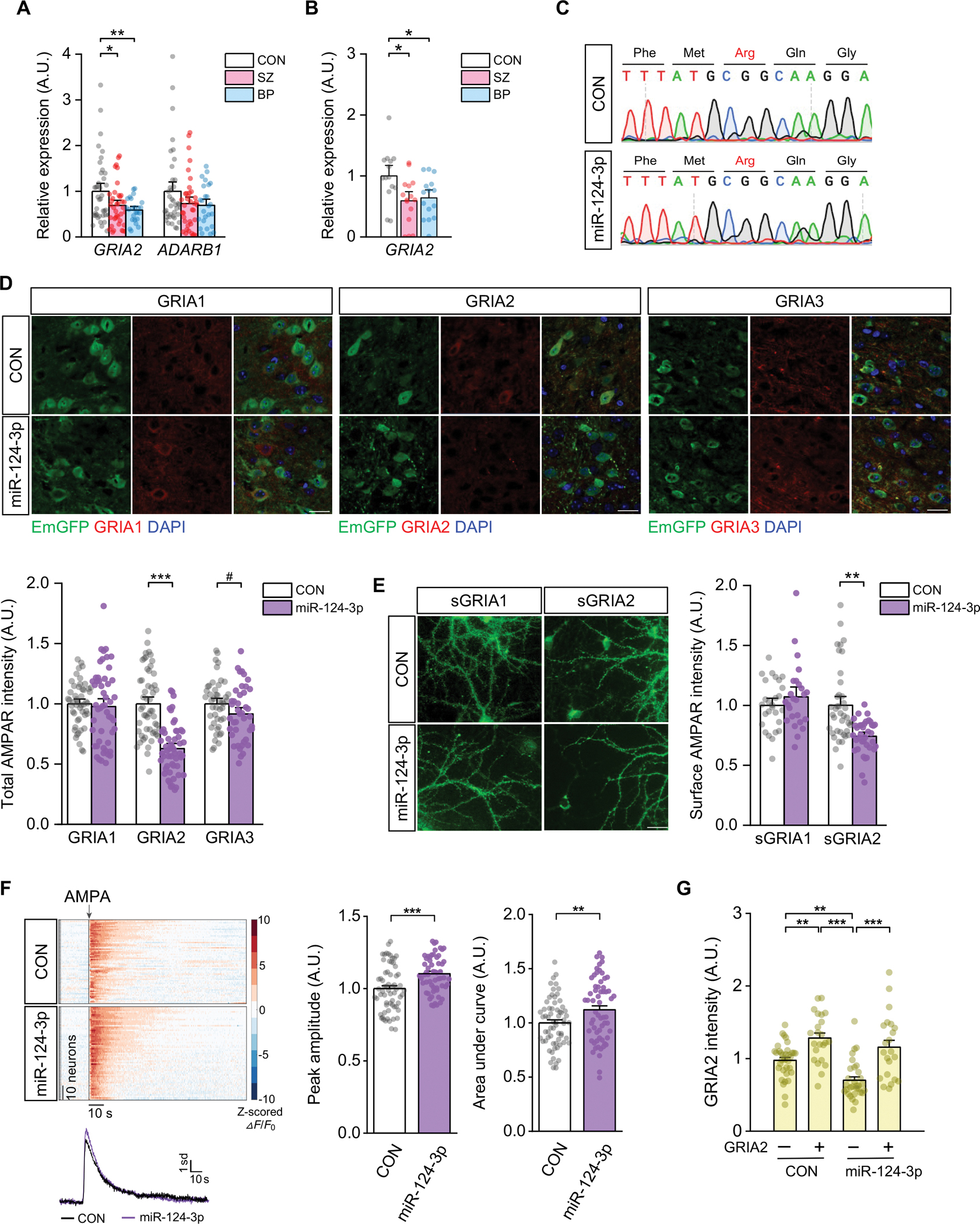
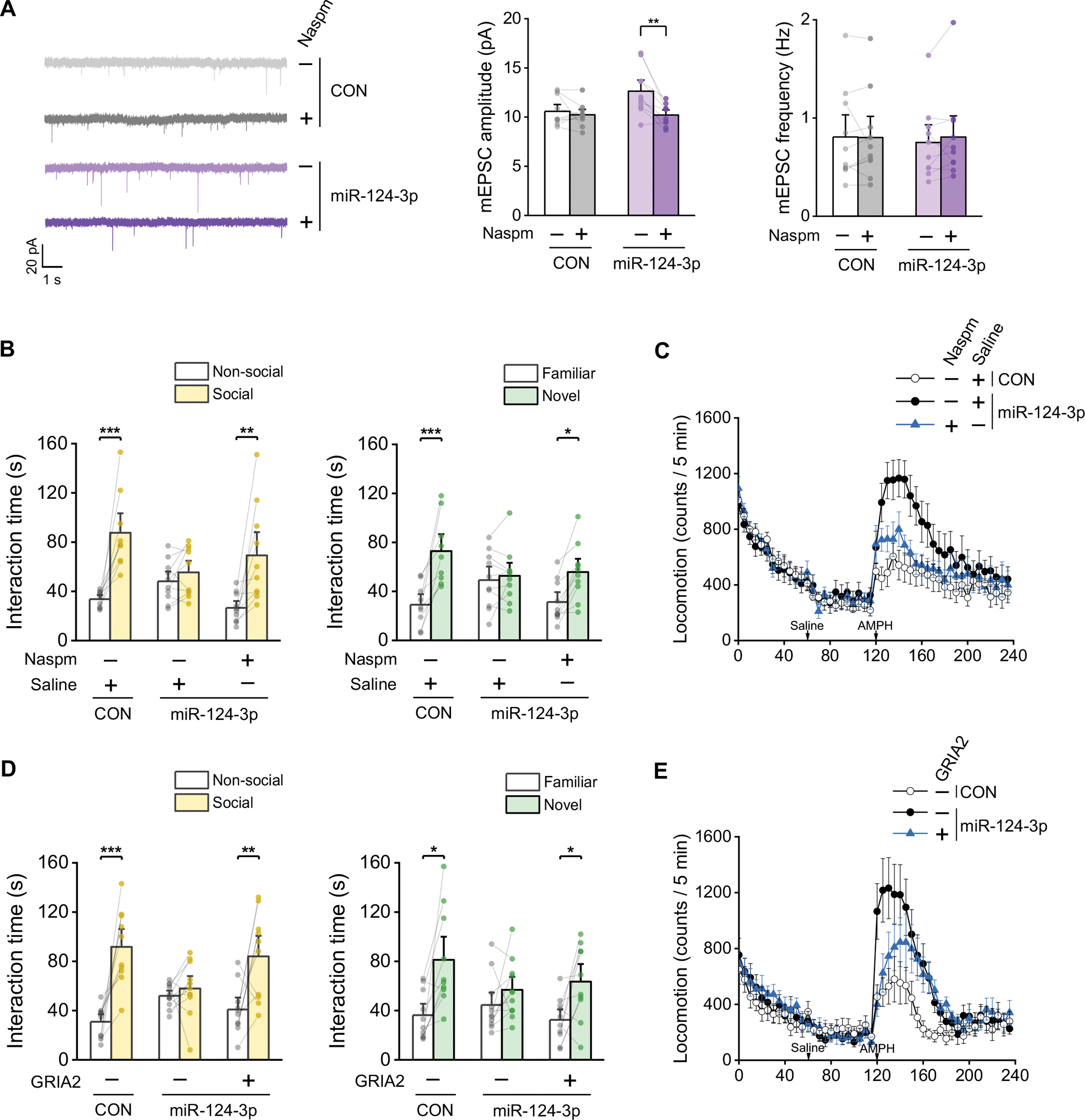
Schizophrenia (SZ) and bipolar disorder (BP) are highly heritable major psychiatric disorders that share a substantial portion of genetic risk as well as their clinical manifestations. This raises a fundamental question of whether, and how, common neurobiological pathways translate their shared polygenic risks into shared clinical manifestations. This study shows the miR-124-3p-AMPAR pathway as a key common neurobiological mediator that connects polygenic risks with behavioral changes shared between these two psychotic disorders. We discovered the upregulation of miR-124-3p in neuronal cells and the postmortem prefrontal cortex from both SZ and BP patients. Intriguingly, the upregulation is associated with the polygenic risks shared between these two disorders. Seeking mechanistic dissection, we generated a mouse model that upregulates miR-124-3p in the medial prefrontal cortex. We demonstrated that the upregulation of miR-124-3p increases GRIA2-lacking calcium-permeable AMPARs and perturbs AMPAR-mediated excitatory synaptic transmission, leading to deficits in the behavioral dimensions shared between SZ and BP.
Keywords: AMPAR; bipolar disorder; miR-124; polygenic risk; schizophrenia.
Copyright © 2022 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests. R.L.H. is a member of the journal’s advisory board.
Figures
Comment in
-
A shared pathway connects schizophrenia and bipolar disorder.Nat Rev Neurosci. 2023 Jan;24(1):2. doi: 10.1038/s41583-022-00662-w. Nat Rev Neurosci. 2023. PMID: 36446901 No abstract available.
-
miR-124-3p mediates polygenic risk shared between schizophrenia and bipolar disorder.Neuron. 2023 Jan 18;111(2):144-146. doi: 10.1016/j.neuron.2022.12.024. Neuron. 2023. PMID: 36657396
References
-
- Consortium T.S.W.G.o.t.P.G., Ripke S, Walters JT, and O’Donovan MC (2020). Mapping genomic loci prioritises genes and implicates synaptic biology in schizophrenia. medRxiv, 2020.2009.2012.20192922. 10.1101/2020.09.12.20192922. - DOI
-
- Mullins N, Forstner AJ, O’Connell KS, Coombes B, Coleman JRI, Qiao Z, Als TD, Bigdeli TB, Børte S, Bryois J, et al. (2021). Genome-wide association study of more than 40,000 bipolar disorder cases provides new insights into the underlying biology. Nat Genet 53, 817–829. 10.1038/s41588-021-00857-4. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
