

Modeling convergent scale-by-scale skin color patterning in multiple species of lizards
- PMID: 36379217
- PMCID: PMC9763091
- DOI: 10.1016/j.cub.2022.10.044
Modeling convergent scale-by-scale skin color patterning in multiple species of lizards
Abstract
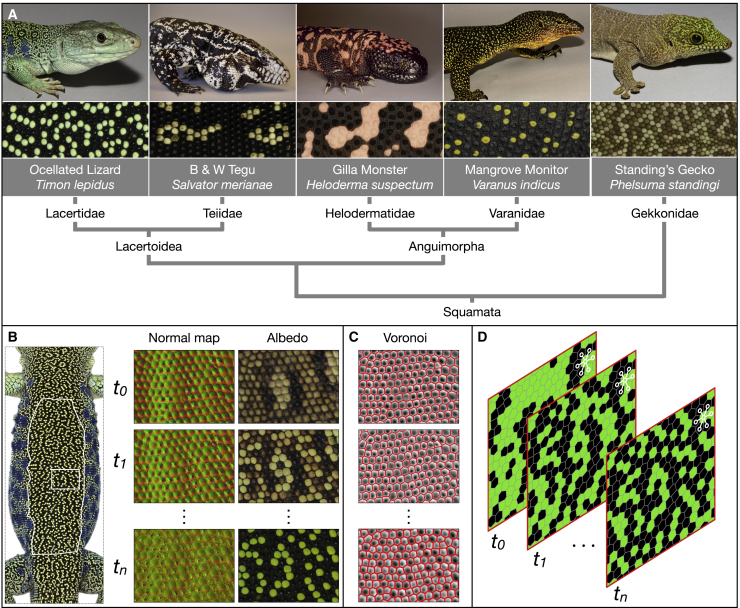
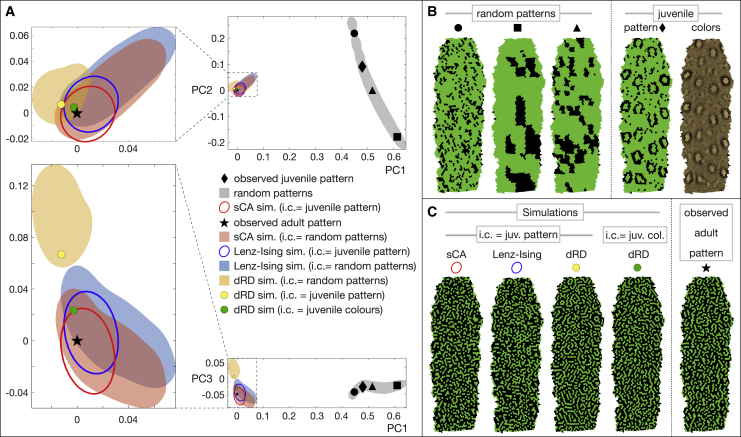
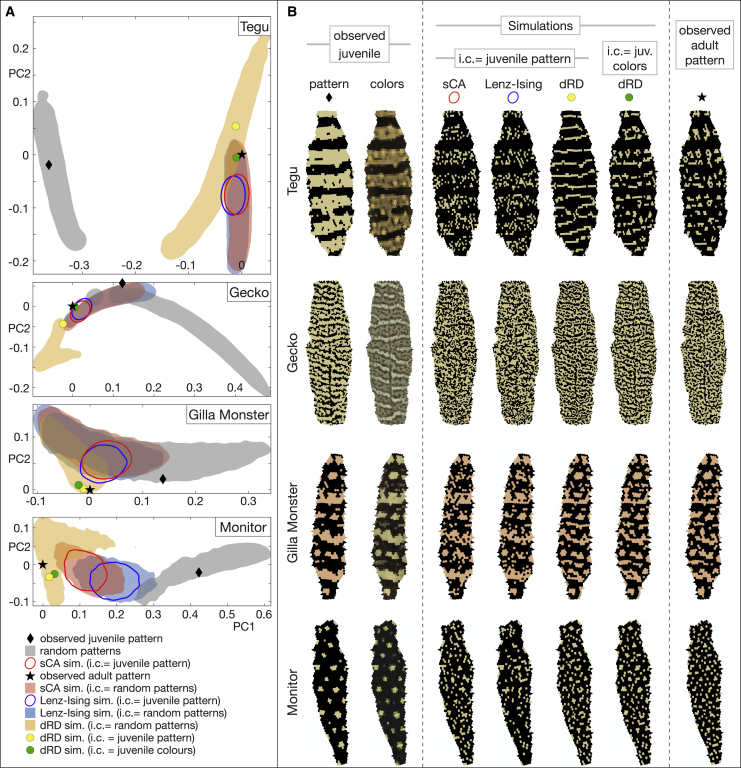
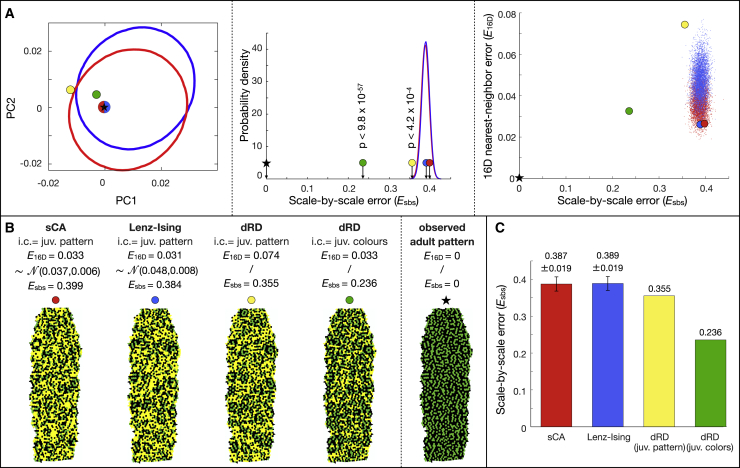
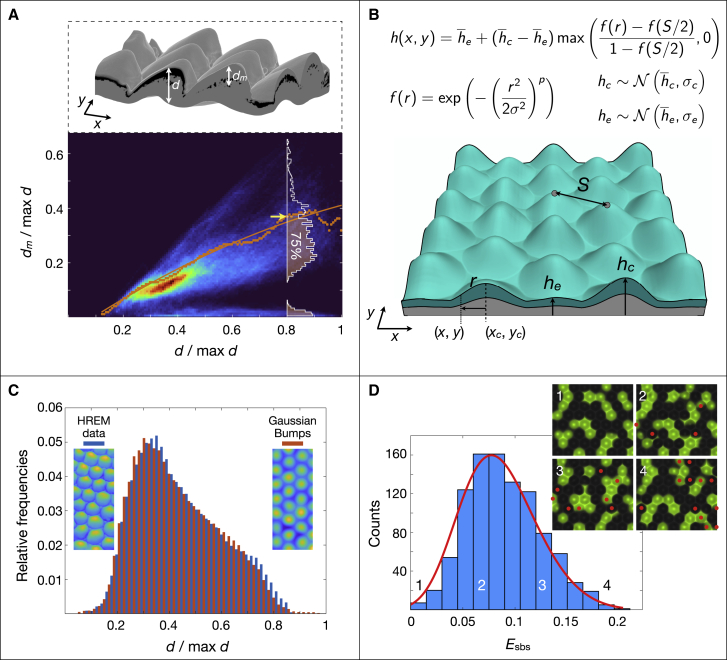
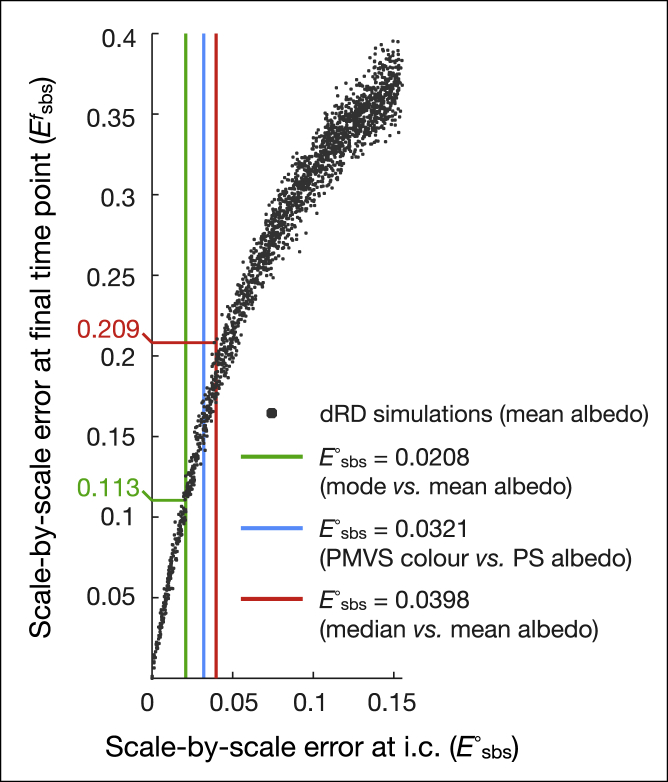
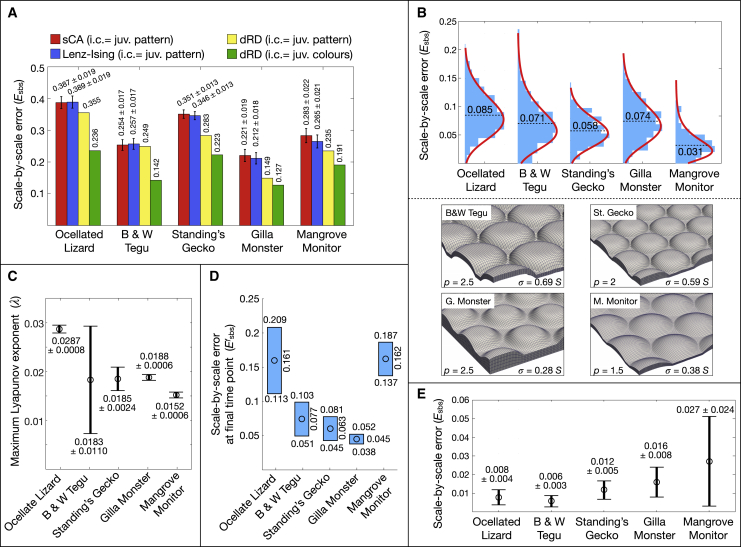
Skin color patterning in vertebrates emerges at the macroscale from microscopic cell-cell interactions among chromatophores. Taking advantage of the convergent scale-by-scale skin color patterning dynamics in five divergent species of lizards, we quantify the respective efficiencies of stochastic (Lenz-Ising and cellular automata, sCA) and deterministic reaction-diffusion (RD) models to predict individual patterns and their statistical attributes. First, we show that all models capture the underlying microscopic system well enough to predict, with similar efficiencies, neighborhood statistics of adult patterns. Second, we show that RD robustly generates, in all species, a substantial gain in scale-by-scale predictability of individual adult patterns without the need to parametrize the system down to its many cellular and molecular variables. Third, using 3D numerical simulations and Lyapunov spectrum analyses, we quantitatively demonstrate that, given the non-linearity of the dynamical system, uncertainties in color measurements at the juvenile stage and in skin geometry variation explain most, if not all, of the residual unpredictability of adult individual scale-by-scale patterns. We suggest that the efficiency of RD is due to its intrinsic ability to exploit mesoscopic information such as continuous scale colors and the relations among growth, scales geometries, and the pattern length scale. Our results indicate that convergent evolution of CA patterning dynamics, leading to dissimilar macroscopic patterns in different species, is facilitated by their spontaneous emergence under a large range of RD parameters, as long as a Turing instability occurs in a skin domain with periodic thickness. VIDEO ABSTRACT.
Keywords: Lenz-Ising model; Turing patterns; cellular automaton; convergence; development; evolution; lizards; reaction-diffusion; scales; skin color patterns.
Copyright © 2022 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
Comment in
-
Colour patterns: Predicting patterns without knowing the details.Curr Biol. 2022 Dec 5;32(23):R1306-R1308. doi: 10.1016/j.cub.2022.10.042. Curr Biol. 2022. PMID: 36473439
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Miscellaneous
