

Modeling the Evolution of Rates of Continuous Trait Evolution
- PMID: 36380474
- PMCID: PMC10276627
- DOI: 10.1093/sysbio/syac068
Modeling the Evolution of Rates of Continuous Trait Evolution
Abstract
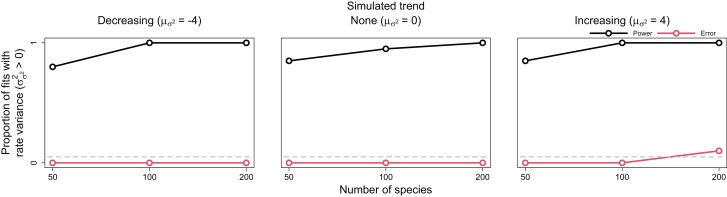
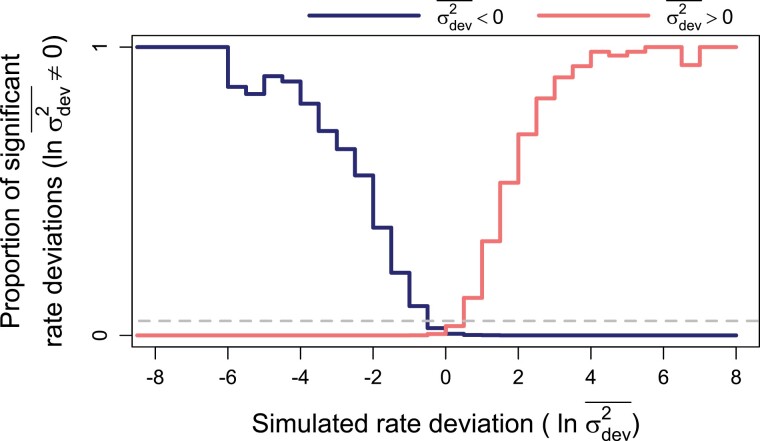
Rates of phenotypic evolution vary markedly across the tree of life, from the accelerated evolution apparent in adaptive radiations to the remarkable evolutionary stasis exhibited by so-called "living fossils." Such rate variation has important consequences for large-scale evolutionary dynamics, generating vast disparities in phenotypic diversity across space, time, and taxa. Despite this, most methods for estimating trait evolution rates assume rates vary deterministically with respect to some variable of interest or change infrequently during a clade's history. These assumptions may cause underfitting of trait evolution models and mislead hypothesis testing. Here, we develop a new trait evolution model that allows rates to vary gradually and stochastically across a clade. Further, we extend this model to accommodate generally decreasing or increasing rates over time, allowing for flexible modeling of "early/late bursts" of trait evolution. We implement a Bayesian method, termed "evolving rates" (evorates for short), to efficiently fit this model to comparative data. Through simulation, we demonstrate that evorates can reliably infer both how and in which lineages trait evolution rates varied during a clade's history. We apply this method to body size evolution in cetaceans, recovering substantial support for an overall slowdown in body size evolution over time with recent bursts among some oceanic dolphins and relative stasis among beaked whales of the genus Mesoplodon. These results unify and expand on previous research, demonstrating the empirical utility of evorates. [cetacea; macroevolution; comparative methods; phenotypic diversity; disparity; early burst; late burst].
© The Author(s) 2022. Published by Oxford University Press on behalf of the Society of Systematic Biologists.
Figures





References
-
- Beaulieu J.M., O’Meara B.C.. 2016. Detecting hidden diversification shifts in models of trait-dependent speciation and extinction. Syst. Biol. 65:583–601. - PubMed
-
- Betancourt M.J., and Girolami M.. 2013. Hamiltonian Monte Carlo for hierarchical models. arXiv 1312.0906.
-
- Blomberg S.P., Garland T. Jr, Ives A.R.. 2003. Testing for phylogenetic signal in comparative data: behavioral traits are more labile. Evolution. 57:717–745. - PubMed
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
