

Elevated CD47 is a hallmark of dysfunctional aged muscle stem cells that can be targeted to augment regeneration
- PMID: 36384141
- PMCID: PMC9746883
- DOI: 10.1016/j.stem.2022.10.009
Elevated CD47 is a hallmark of dysfunctional aged muscle stem cells that can be targeted to augment regeneration
Abstract
In aging, skeletal muscle strength and regenerative capacity decline, due in part to functional impairment of muscle stem cells (MuSCs), yet the underlying mechanisms remain elusive. Here, we capitalize on mass cytometry to identify high CD47 expression as a hallmark of dysfunctional MuSCs (CD47hi) with impaired regenerative capacity that predominate with aging. The prevalent CD47hi MuSC subset suppresses the residual functional CD47lo MuSC subset through a paracrine signaling loop, leading to impaired proliferation. We uncover that elevated CD47 levels on aged MuSCs result from increased U1 snRNA expression, which disrupts alternative polyadenylation. The deficit in aged MuSC function in regeneration can be overcome either by morpholino-mediated blockade of CD47 alternative polyadenylation or antibody blockade of thrombospondin-1/CD47 signaling, leading to improved regeneration in aged mice, with therapeutic implications. Our findings highlight a previously unrecognized age-dependent alteration in CD47 levels and function in MuSCs, which underlies reduced muscle repair in aging.
Keywords: CyTOF; U1 snRNA; aging; alternative polyadenylation; antisense morpholino oligonucleotide; in vivo antibody blockade; muscle stem cells; paracrine signaling; regeneration; thrombospondin-1/CD47 signaling.
Copyright © 2022 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests H.M.B. is cofounder of Rejuvenation Technologies Inc. and Epirium Bio. H.M.B. and E.P. are named inventors on patent application no. PCT/US2021/038549 held by Stanford University.
Figures
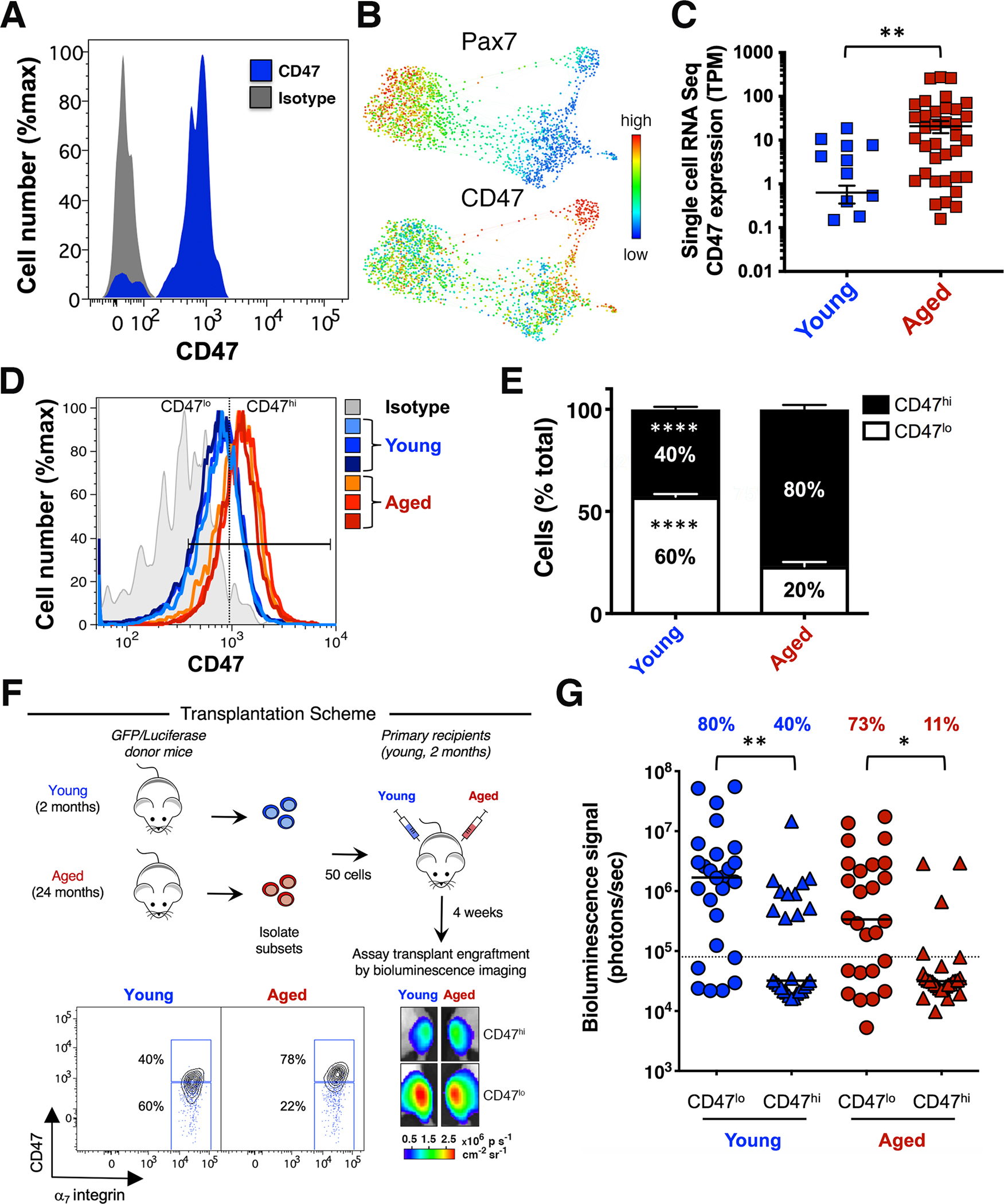
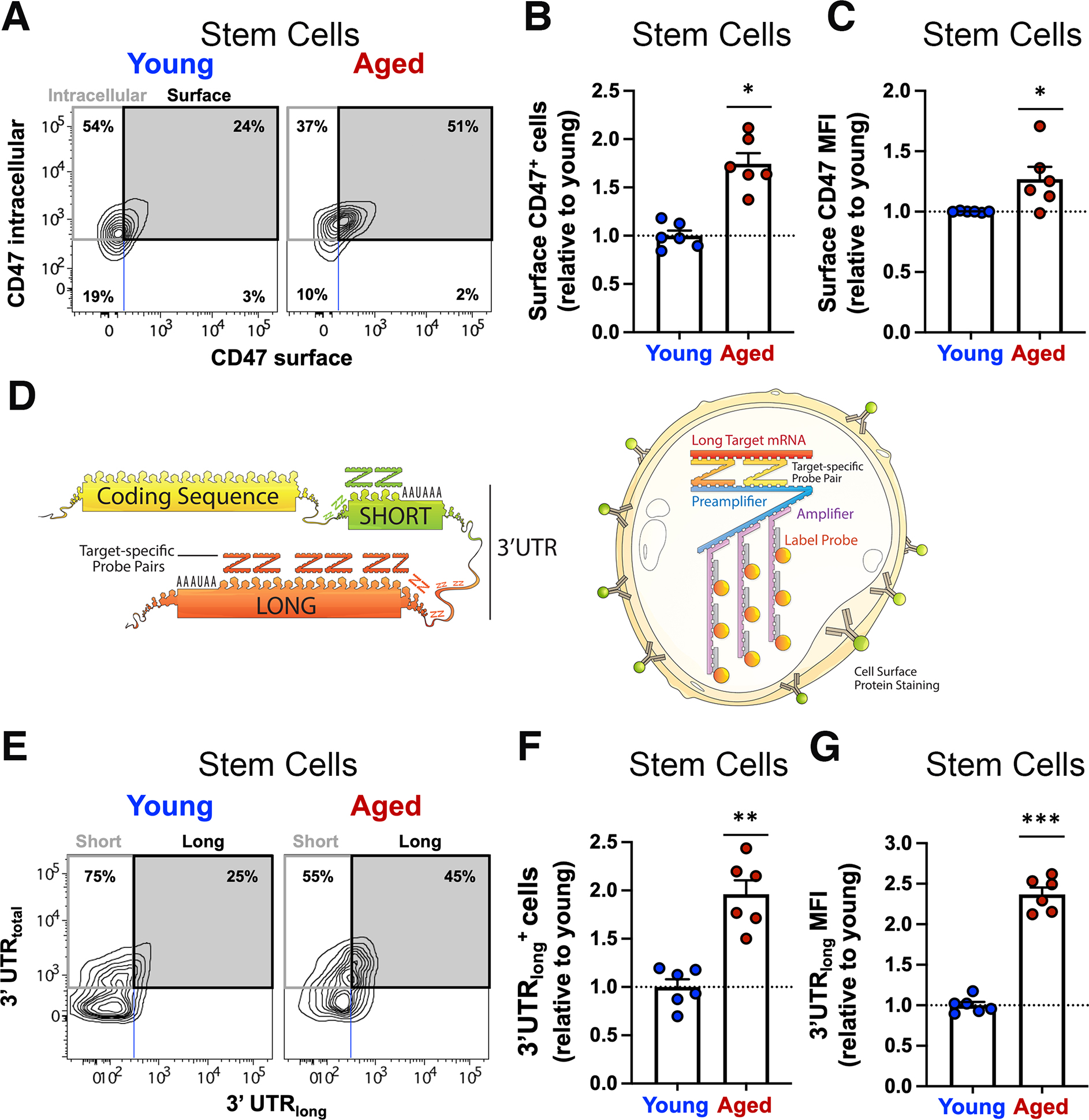

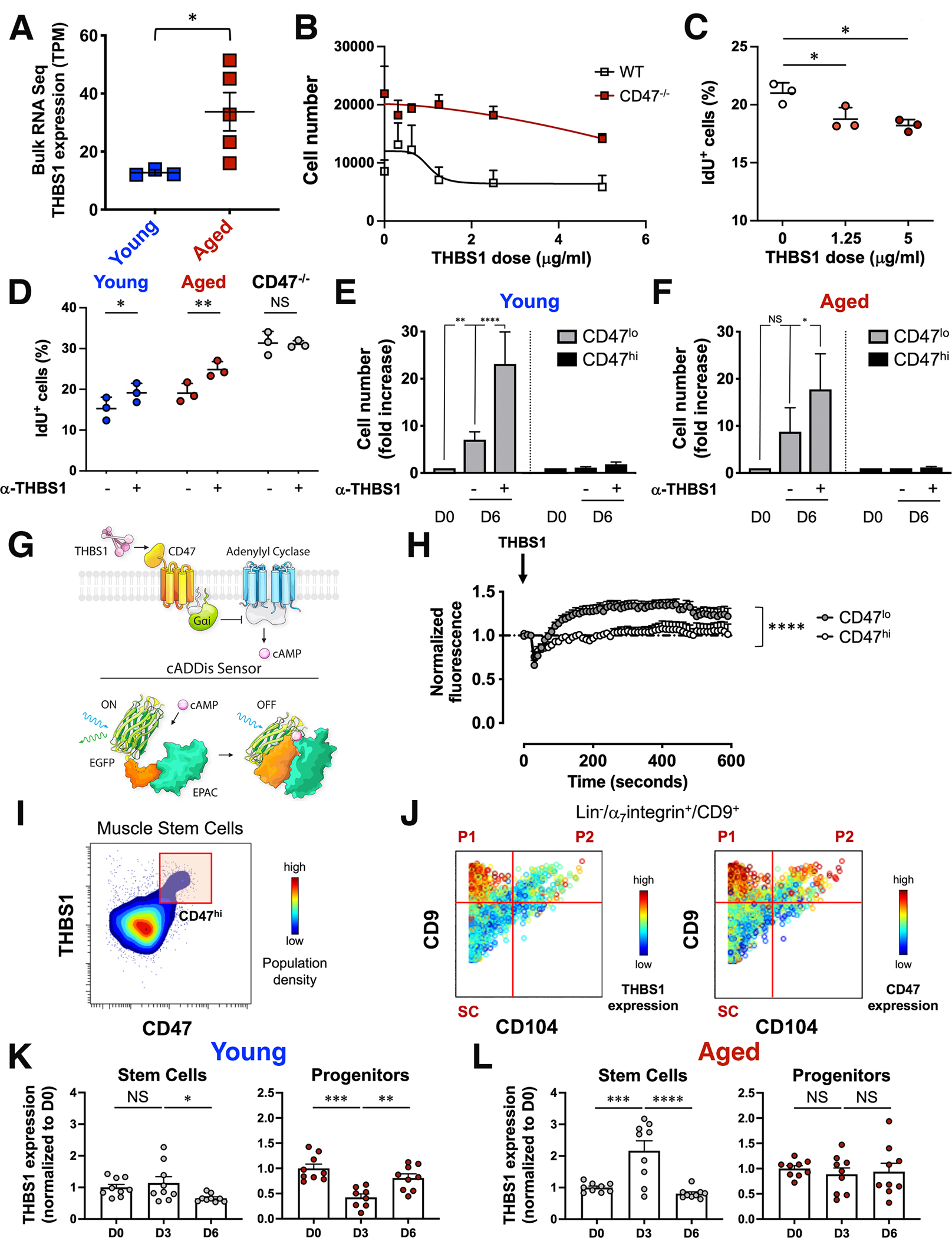


Comment in
-
A post-middle-age crisis for CD47 and THBS1 that turns into a vicious cycle.Cell Stem Cell. 2022 Dec 1;29(12):1613-1615. doi: 10.1016/j.stem.2022.11.003. Cell Stem Cell. 2022. PMID: 36459961
Similar articles
-
Small molecule nicotinamide N-methyltransferase inhibitor activates senescent muscle stem cells and improves regenerative capacity of aged skeletal muscle.Biochem Pharmacol. 2019 May;163:481-492. doi: 10.1016/j.bcp.2019.02.008. Epub 2019 Feb 10. Biochem Pharmacol. 2019. PMID: 30753815 Free PMC article.
-
Relayed signaling between mesenchymal progenitors and muscle stem cells ensures adaptive stem cell response to increased mechanical load.Cell Stem Cell. 2022 Feb 3;29(2):265-280.e6. doi: 10.1016/j.stem.2021.11.003. Epub 2021 Dec 1. Cell Stem Cell. 2022. PMID: 34856120
-
Primary cilia on muscle stem cells are critical to maintain regenerative capacity and are lost during aging.Nat Commun. 2022 Mar 17;13(1):1439. doi: 10.1038/s41467-022-29150-6. Nat Commun. 2022. PMID: 35301320 Free PMC article.
-
Stem Cell Aging in Skeletal Muscle Regeneration and Disease.Int J Mol Sci. 2020 Mar 6;21(5):1830. doi: 10.3390/ijms21051830. Int J Mol Sci. 2020. PMID: 32155842 Free PMC article. Review.
-
Decline of regenerative potential of old muscle stem cells: contribution to muscle aging.FEBS J. 2023 Mar;290(5):1267-1289. doi: 10.1111/febs.16352. Epub 2022 Jan 25. FEBS J. 2023. PMID: 35029021 Review.
Cited by
-
Beyond cancer: The potential application of CD47-based therapy in non-cancer diseases.Acta Pharm Sin B. 2025 Feb;15(2):757-791. doi: 10.1016/j.apsb.2024.11.018. Epub 2024 Nov 28. Acta Pharm Sin B. 2025. PMID: 40177549 Free PMC article. Review.
-
CD47-blocking antibody confers metabolic benefits against obesity.Cell Rep Med. 2025 May 20;6(5):102089. doi: 10.1016/j.xcrm.2025.102089. Epub 2025 Apr 22. Cell Rep Med. 2025. PMID: 40267910 Free PMC article.
-
Structural-functional diversity of CD47 proteoforms.Front Immunol. 2024 Feb 15;15:1329562. doi: 10.3389/fimmu.2024.1329562. eCollection 2024. Front Immunol. 2024. PMID: 38426113 Free PMC article. Review.
-
The role of CD47 in non-neoplastic diseases.Heliyon. 2023 Nov 29;9(12):e22905. doi: 10.1016/j.heliyon.2023.e22905. eCollection 2023 Dec. Heliyon. 2023. PMID: 38125492 Free PMC article. Review.
-
Mini-MEndR: a miniaturized 96-well predictive assay to evaluate muscle stem cell-mediated repair.BMC Methods. 2024;1(1):5. doi: 10.1186/s44330-024-00005-4. Epub 2024 Jun 7. BMC Methods. 2024. PMID: 38872952 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
