

Impact of nutrients on the function of the chlamydial Rsb partner switching mechanism
- PMID: 36385643
- PMCID: PMC12173453
- DOI: 10.1093/femspd/ftac044
Impact of nutrients on the function of the chlamydial Rsb partner switching mechanism
Abstract
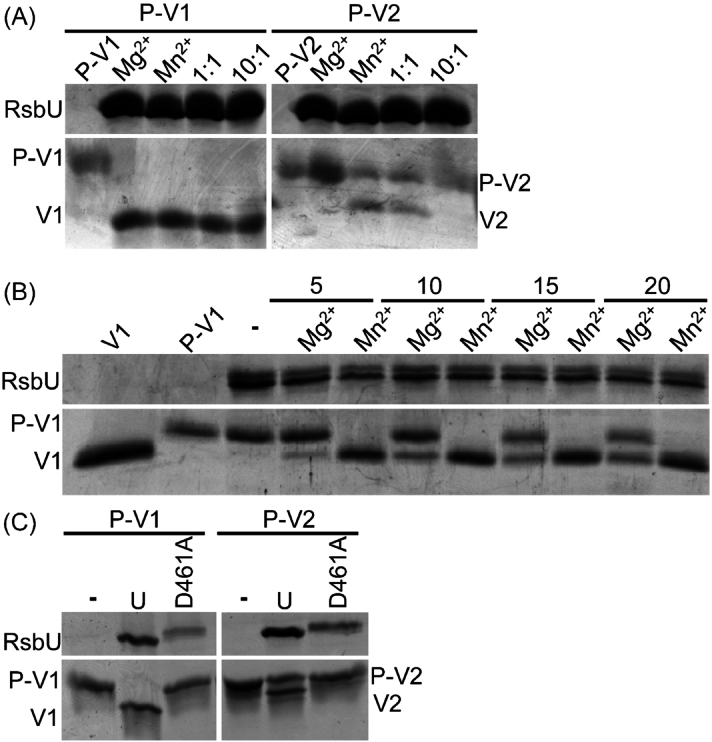
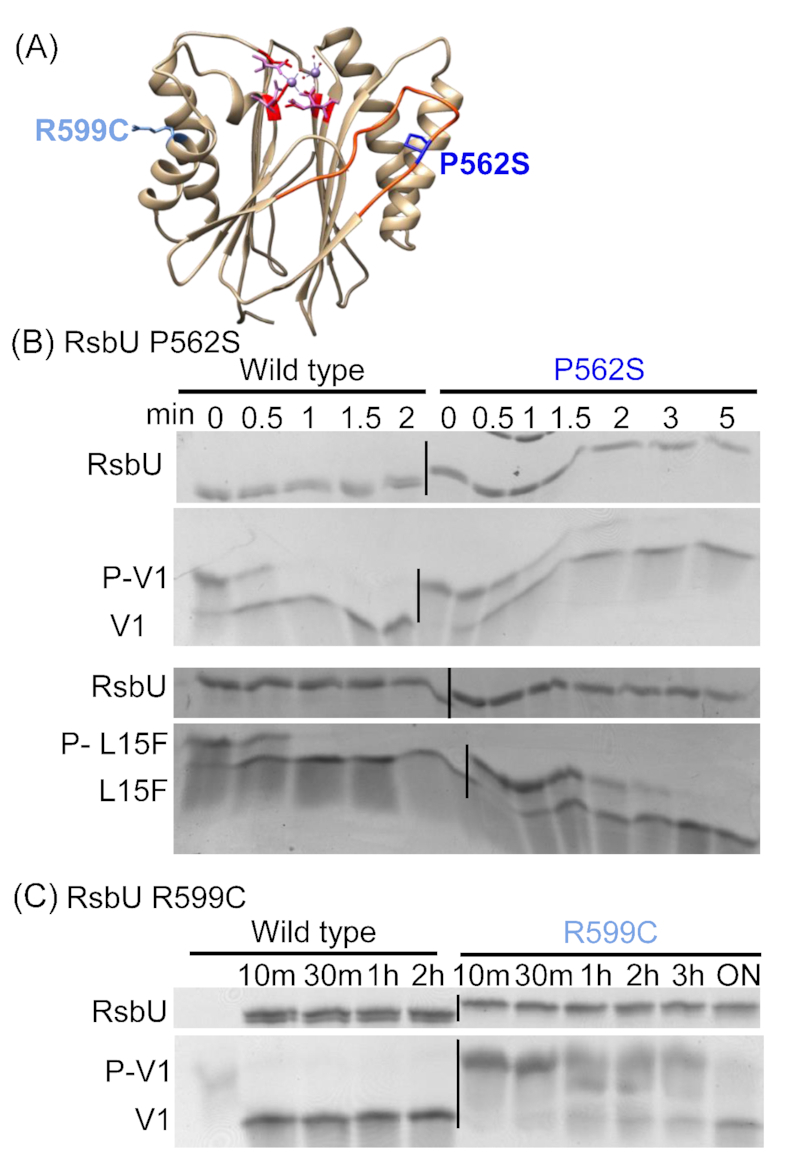
The obligate intracellular bacterial pathogen Chlamydia trachomatis is a leading cause of sexually transmitted infections and infectious blindness. Chlamydia undergo a biphasic developmental cycle alternating between the infectious elementary body (EB) and the replicative reticulate body (RB). The molecular mechanisms governing RB growth and RB-EB differentiation are unclear. We hypothesize that the bacterium senses host cell and bacterial energy levels and metabolites to ensure that development and growth coincide with nutrient availability. We predict that a partner switching mechanism (PSM) plays a key role in the sensing and response process acting as a molecular throttle sensitive to metabolite levels. Using purified wild type and mutant PSM proteins, we discovered that metal type impacts enzyme activity and the substrate specificity of RsbU and that RsbW prefers ATP over GTP as a phosphate donor. Immunoblotting analysis of RsbV1/V2 demonstrated the presence of both proteins beyond 20 hours post infection and we observed that an RsbV1-null strain has a developmental delay and exhibits differential growth attenuation in response to glucose levels. Collectively, our data support that the PSM regulates growth in response to metabolites and further defines biochemical features governing PSM-component interactions which could help in the development of novel PSM-targeted therapeutics.
Keywords: Chlamydia; PP2C; Rsb; glucose; metalation; phosphorylation.
© The Author(s) 2022. Published by Oxford University Press on behalf of FEMS.
Conflict of interest statement
The authors declare no competing financial interest.
Figures




Similar articles
-
Distinct impacts of each anti-anti-sigma factor ortholog of the chlamydial Rsb partner switching mechanism on development in Chlamydia trachomatis.Microbiol Spectr. 2024 Oct 29;12(12):e0184624. doi: 10.1128/spectrum.01846-24. Online ahead of print. Microbiol Spectr. 2024. PMID: 39470281 Free PMC article.
-
Tag-Dependent Substrate Selection of ClpX Underlies Secondary Differentiation of Chlamydia trachomatis.mBio. 2022 Oct 26;13(5):e0185822. doi: 10.1128/mbio.01858-22. Epub 2022 Sep 26. mBio. 2022. PMID: 36154190 Free PMC article.
-
Light and polyphosphate kinase 2 cooperatively regulate the production of zero-valent sulfur in a deep-sea bacterium.mSystems. 2025 Jun 17;10(6):e0047325. doi: 10.1128/msystems.00473-25. Epub 2025 May 16. mSystems. 2025. PMID: 40377319 Free PMC article.
-
Pathogenicity and virulence of Chlamydia trachomatis: Insights into host interactions, immune evasion, and intracellular survival.Virulence. 2025 Dec;16(1):2503423. doi: 10.1080/21505594.2025.2503423. Epub 2025 May 15. Virulence. 2025. PMID: 40353442 Free PMC article. Review.
-
Assessing the comparative effects of interventions in COPD: a tutorial on network meta-analysis for clinicians.Respir Res. 2024 Dec 21;25(1):438. doi: 10.1186/s12931-024-03056-x. Respir Res. 2024. PMID: 39709425 Free PMC article. Review.
Cited by
-
Distinct impacts of each anti-anti-sigma factor ortholog of the chlamydial Rsb partner switching mechanism on development in Chlamydia trachomatis.Microbiol Spectr. 2024 Oct 29;12(12):e0184624. doi: 10.1128/spectrum.01846-24. Online ahead of print. Microbiol Spectr. 2024. PMID: 39470281 Free PMC article.
References
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
